
 被子植物
被子植物簡介
被子植物分為兩個綱----雙子葉植物綱(木蘭綱)和單子葉植物綱(百合綱)。 被子植物拉丁名 Angiospermae,來源於希臘文複合詞。Angio—包被的與—sperma種子,兩者接合,用複數第一格即成 Angiospermae,英文名為 Angiosperm。
 被子植物
被子植物極端的自然環境還有沙漠。如中國新疆省的沙漠地區,有胡楊(Populus diversifolia )和梭梭(Haloxylon ammodendron)生存,能適應乾旱氣候。北非撒哈拉大沙漠中下雨極少,有的地方十幾年無雨。有一種植物叫矮生齒子草 (Odontospermum pygmaeum),由於極端乾旱形成只有幾十天極短的生命周期,稱為短命植物。它在稍有雨水時,能發芽生長到開花結實,完成一代任務。平時稍有濕潤,花就張開,一旦乾燥,花即閉合,十分靈敏。美洲墨西哥的沙漠地區,有一類特殊適應乾旱的植物就是多漿植物,著名的為仙人掌科。全身多刺,葉退化,莖含水多用以抗旱。其中有的種類形如巨人,如用刀砍開,可以直接喝到水。在鹽鹼地上,有抗鹽性強的被子植物,以黎科最著名。如鹽角草(Salicornia herbacea)為一年生草本,肉質,葉極小。莖節狀,可以行光合作用。在它的莖的橫切面上有引人注目的特徵:如表皮薄而光滑,柵欄組織有兩層,內部細胞大、含水多,維管組織在中心。多水性是由於鈉(Na)離子影響下形成的。鹽角草活物重量的92%以上為水分。乾燥了的鹽角草燃燒後留下的灰分極多,占乾量的45%以上。上述的莖結構、含水多和灰分多,均是典型的鹽生植物特性。鹽角草生在鹽沼地上,是鹽分最多的地方,可為其標準代表。黎科的鹽生植物的進化就是在鹽演條件下進行的。
起源
1.起源時間
當前多數學者認為被子植物起源於白堊紀或晚侏羅紀。根據斯科特(Scott)、馬朗(Ba-rghoon)和利奧波德(Leopold)(1960)對以前記述過的化石進行了全面的討論,發現白堊紀之前未曾保存具確實證據的被子植物化石。此外從孢粉證據來看,同樣表現在白堊紀以前的地層中,未能找到被子植物花粉。多伊爾(Doyle 1969)和
 被子植物
被子植物2.發源地
關於被子植物的發源地存在著十分對立的觀點:即高緯度——北極或南極起源說和低緯度——熱帶或亞熱帶起源說。目前,大多數學者支持後一個學說。近數十年來的資料表明,大量被子植物化石在中、低緯度出現的時間實際上早於高緯度。被子植物在熱帶或亞熱帶首先出現,然後逐漸向高緯度地區擴展。
現代被子植物地理分布情況,同樣說明被子植物可能起源於中、低緯度地區。在被子植物現存的400餘科中,有半數以上的科依然集中分布在中、低緯度地區,尤其是那些較原始的木蘭科、八角科、昆欄樹科和水青樹科等更是如此。貝利(Bailey 1949)、史密斯(Smith 1967)和塔赫他間(Takhtajan 1969)以現代植物科的分布及化石證據的分析,發現西南太平洋和東南亞地區原始毛茛類型(廣義的木蘭目)分布占優勢,認為這個地區可能是被子植物早期分化的地區。中國的吳征縊(1964,1977)認為中國南部、西南部和中南半島在北緯20º~40º間的廣大地區最富於特有的古老科屬,可能是被子植物的發源地。Camp (1952)提出南美亞馬遜河流域的平原地區熱帶雨林中有許多接近原始類型的被子植物,被子植物可能起源於這一區域。
總之,被子植物起源的地區仍處於推測階段,由於化石植物的缺乏和對過去發生的地質、氣候變化還不十分清楚,雖多數學者贊同低緯度起源,但確切回答被子植物的起源地點是有困難的,還有待今後更深入的研究。
3.可能的祖先
被子植物的屬種十分龐雜,形態變化很大,分布極廣,粗看起來,確實難用統一的特徵將所有的被子植物歸成一類。因此,對被子植物的祖先存在不同的假說,有多元論和單元論兩種起源說。
多元論認為被子植物來自許多不相親近的群類,彼此是平行發展的。胡先驌、米塞(Meeuse)、恩格勒(Engler)和蘭姆(Lam)等人是多元論的代表。中國的分類學家胡先驌1950年發表了一個被子植物多元起源的系統,也是中國學者發表的被子植物的唯一系統。
單元論是目前多數植物學家主張的被子植物起源說。主要依據是被子植物有許多獨特和高度特
化的性狀,如雄蕊都有四個孢子(花粉)囊和特有的藥室內層;大孢子葉(心皮)和柱頭的存在;雌雄蕊在花軸排列的位置固定不變;雙受精現象和三倍體胚乳;以及篩管和伴胞的存在。因此,人們認為被子植物只能起源於一個共同的祖先。哈欽森(Hutchinson)、塔赫他間(Takhtajan)、克朗奎斯特(Cronquist)和 Judd是單元論的主要代表。
被子植物如確係單元起源,那么它究竟發生於哪一類植物呢?推測很多,至今未有定論。人們推測的有:藻類、蕨類、松杉目、買麻藤目、本內蘇鐵目、種子蕨等。目前比較流行的是本內蘇鐵目和種子蕨這二種假說。
 被子植物
被子植物4.被子植物系統發育重建
人類研究植物分類、進化及系統發育的目的是要揭示植物世界發生、發展、絕滅的歷史,從現存被子植物和化石記錄,通過巨觀與微觀特徵的研究重建被子植物的系統發育,建立一個完全自然的系統。
特徵
被子植物是植物界進化最高級、種類最多、分布最廣的類群。現知被子植物有 l萬多屬,20多萬種,占植物界的一半。中國有2700多屬,約3萬種,是藥用植物最多的類群。被子植物的種類能有如此眾多,適應性有如此廣泛,這和它的結構複雜化、完善化分不開的,特別是繁殖器官的結構和生殖過程的特點,給予了它適應、抵禦各種不良環境的內在條件,使它在生存競爭、自然選擇的矛盾鬥爭過程中不斷產生新的變異,產生新的物種,而在地球上占絕對優勢。和裸子植物相比,被子植物有真正的花,故又叫有花植物;胚珠包藏在子房內,得到良好的保護,子房在受精後形成的果實既保護種子又以各種方式幫助種子散布;具有雙受精現象和三倍體的胚乳,此種胚乳不是單純的雌配子體,而具有雙親的特性,使新植物體有更強的生活力;孢子體高度發達和進一步分化,除喬木和灌木外,更多是草本;在解剖構造上,木質部中有導管,韌皮部有篩管、伴胞,使輸導組織結構和生理功能更加完善,同時在化學成份上,隨著被子植物的演化而不斷發展和複雜化,被子植物包含了所有天然化合物的各種類型,具有多種生理活性。列舉的被子植物的五個進化特徵,是與裸子植物相比較而得出的,能產生種子、精子靠花粉管傳送、有胚乳等種子植物共有的特徵。
(一)具有真正的花
典型的被子植物的花由花尊、花冠、雄蕊群、雌蕊群4部分組成,各個部分稱為花部。被子植物花的各部在數量上、形態上有極其多樣的變化,這些變化是在進化過程中, 適應於蟲媒、風媒、鳥媒、或水媒傳粉的條件,被自然界選擇,得到保留,並不斷加強造成的。
(二)具雌蕊
雌蕊由心皮所組成,包括子房、花柱和柱頭3部分。胚珠包藏在子房內,得到子房的保護, 避免了昆蟲的咬噬和水分的喪失。子房在受精後發育成為果實。果實具有不同的色、香、味,多種開裂方式;果皮上常具有各種鉤、刺、翅、毛。果實的所有這些特點,對於保護種子成熟,幫助種子散布起著重要作用,它們的進化意義也是不言而喻的。
(三)具有雙受精現象
雙受精現象,即兩個精細胞進入胚囊以後,1個與卵細胞結合形成合子,另1個與2個極核結合,形成3n染色體,發育為胚乳,幼胚以3n染色體的胚乳為營養,使新植物體內矛盾增大,因而具有更強的生活力。所有被子植物都有雙受精現象,這也是它們有共同祖先的一個證據。
(四)孢子體高度發達被子植物的孢子體,在形態、結構、生活型等方面,比其他各類植物更完善化、多樣化,有世界上最高大的喬木,如杏仁按(Eucalyptus amygdalina Labill.),高達156米;也有微細如沙粒的小草本如無根萍[Wolffia arrhiza (L.) Wimm.],每平方米水面可容納300萬個個體 有重達25千克僅含1顆種子的果實,如王棕(大王椰子)[Roystonea regia (H.B.K.) O. F.Cook];也有輕如塵埃,5萬顆種子僅重0.1克的植物如熱帶雨林中的一些附生蘭;有壽命長達6千年的植物,如龍血樹(Dracaena draco L.);也有在3周內開花結籽完成生命周期的植物(如一些生長在荒漠的十字花科植物);有水生、砂生、石生和鹽鹼地生的植物;有自養的植物也有腐生、寄生的植物。在解剖構造上,被子植物的次生木質部有導管,韌皮部有伴胞;而裸子植物中一般均為管胞(只有麻黃和買麻藤類例外),韌皮部無伴胞,輸導組織的完善使體內物質運輸暢通,適應性得到加強。
(五)配子體進一步退化(簡化)
被子植物的小孢子(單核花粉粒)發育為雄配子體,大部分成熟的雄配子體僅具2個細胞(2核花粉粒),其中1個為營養細胞,I個為生殖細胞,少數植物在傳粉前生殖細胞就分裂1次,產生2個精子,所以這類植物的雄配子體為3核的花粉粒,如石竹亞綱的植物和油菜、玉米、大麥、小麥等。被子植物的大孢子發育為成熟的雌配子體稱為胚囊,通常胚囊只有8個細胞:3個反足細胞、2個極核、2個助細胞、1個卵。反足細胞是原葉體營養部分的殘餘。有的植物(如竹類)反足細胞可多達300餘個,有的(如蘋果、梨)在胚囊成熟時,反足細胞消失。助細胞和卵合稱卵器,是頸卵器的殘餘。由此可見,被子植物的雌、雄配子體均無獨立生活能力,終生寄生在孢子體上,結構上比裸子植物更簡化。配子體的簡化在生物學上具有進化的意義。
被子植物的上述特徵,使它具備了在生存競爭中,優越於其他各類植物的內部條件。被子植物的產生,便地球上第一次出現色彩鮮艷、類型繁多、花果豐茂的景象,隨著被子植物花的形態的發展,果實和種子中高能量產物的貯存,使得直接或間接地依賴植物為生的動物界(尤其是昆蟲、鳥類和哺乳類),獲得了相應的發展,迅速地繁茂起來。
分類
 被子植物
被子植物雙子葉植物綱 單子葉植物綱
根 直根系 鬚根系
莖 維管柬成環狀排列,有形成層 維管束成星散排列,無形成層
葉 具網狀脈 具平形脈或弧形脈
花 各部分基數為4或5 各部分基數為3
花粉粒具3個萌發孔 花粉粒具單個萌發孔
胚 具2片子葉 具1片子葉
以上區別點不是絕對的,實際上有交錯現象,如雙子葉植物綱中的毛茛科、車前科、菊科等有鬚根系植物;胡椒科、睡蓮科、毛茛科、石竹科等有維管束星散排列的植物;樟科、木蘭科、小檗科、毛茛科有3基數的花;睡蓮科、毛茛科、小續科、罌粟科、傘形科等有 l片子葉的現象。單子葉植物綱中的天南星科、百合科、薯蕷科等有網狀脈;眼子菜科、百合科、百部科等有4基數的花
雙子葉植物綱 被子植物
被子植物離瓣花亞綱又稱古生花被亞綱 (Archichlamydeae)。包括無被花,單被花或有花萼和花冠區別,而花瓣通常分離的類型。雄蕊和花冠離生。胚殊一般有一層珠被。
離瓣花亞綱恩格勒系統分科目錄
(二)合瓣花亞綱 Sympetalae
本亞綱又稱後生花被亞綱(Metachlamydeae),主要特徵是花瓣多少連合成合瓣花冠。花冠形成了各種形狀,如漏斗狀、鍾狀、唇形、管狀、舌狀等,由輻射對稱發展到兩側對稱。花冠各式的連合,增加了對昆蟲傳粉的適應及對雄蕊和雌蕊的保護。因此,合瓣花類群比離瓣花類群進化。
花的輪數趨向減少,由5輪(花萼 l輪,花瓣 l輪,雄蕊2輪,雌蕊的心皮 l輪,如杜鵑花科、柿樹科等)減為4輪(花萼、花瓣、雄蕊、心皮均為 l輪,如木犀科、忍冬科等),各輪數目也逐步減少,如雄蕊的數目從與花冠裂片同數,如由5(鏇花科、茄科、報春花科)減為4—2(唇形科、葫蘆科),心皮數由5(杜鵑花科)減為2(四輪花類的心皮均多為2個)。
通常無托葉,胚珠只有一層胚被。
分類系統
19世紀以來,許多植物分類工作者為建立一個“自然”的分類系統作出了巨大努力。他們根據各自的系統發育理論,提出的分類系統已有數十個。但由於有關被子植物起源、演化的知識特別是化石證據不足,直到現在還沒有一個比較完善的分類系統。目前世界上運用比較廣泛的仍是恩格勒系統和哈欽松系統。在各級分類系統的安排上,克朗奎斯特系統和塔赫他間系統被認為更為合理。
一、恩格勒系統
這是德國分類學家思格勒(A.Engler)和勃蘭特(K.PrantI)於1897年在其《植物自然分科志》巨著中所使用的系統,它是分類學史上第一個比較完整的系統,它將植物界分13門,第13門為種子植物門,再分為裸子植物和被子植物兩個亞門,被子植物亞門包括單子葉植物和雙子葉植物兩個綱,並將雙子葉植物綱分為離瓣花亞綱(古生花被亞綱)和合瓣花亞綱(後生花被亞綱)。
恩格勒系統將單子葉植物放在雙子葉植物之前,將合瓣花植物歸併一類,認為是進化的一群植物,將柔荑花序植物作為雙子葉植物中最原始的類群,而把木蘭目、毛茛目等認為是較為進化的類群,這些觀點為現代許多分類學家所不贊同。
思格勒系統幾經修訂,在1964年出版的《植物分科志要》第十二版中,已把雙子葉植物放在單子葉植物之前。共有62目,344科,其中雙子葉植物48目,290科,單子葉植物14目,54科。藥用植物學教材被子植物分類部分採用修訂的思格勒系統,但有的內容有變動。
二、哈欽松系統
這是英國植物學家哈欽松(J.Hutchinson)於1926年和1934年在其《有花植物科志》I、II中所建立的系統。在1973年修訂的第三版中,共有111目,411科,其中雙子葉植物82目,342科,單子葉植物29目,69科。
哈欽松系統認為多心皮的木蘭目、毛茛目是被子植物的原始類群,但過分強調了木本和草本兩個來源,認為木本植物均由木蘭目演化而來,草本植物均由毛茛目演化而來,結果使得親緣關係很近的一些科在系統位置上都相隔很遠,如草本的傘形科和木本的山茱萸科、五加科;草本的唇形科和木本的馬鞭草科等,這種觀點亦受到現代多數分類學家所反對。
三、塔赫他間系統
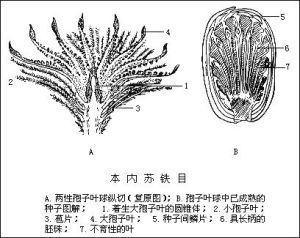 被子植物
被子植物塔赫他間系統經過多次修訂,在1980年修訂版中,共有28超目,92目,416科,其中雙予葉植物(木蘭綱)20超目,71目333科,單子葉植物(百合綱)8超目,21目,77科,顯得較繁鎖。
四、克朗奎斯特系統
這是美國植物學家克朗奎斯特(A.Cronquist)於1968年在其《有花植物的分類和演化》一書中發表的系統。在1981年修訂版中,共有83目,388科,其中雙子葉植物64目,318科,單幹葉植物19目,65科。
克朗奎斯特系接近於塔赫他間系統,把被子植物門(稱木蘭植物門)分成木蘭綱和百合綱,但取消了“超目”一級分類單元,科的劃分也少於塔赫他間系統。現中國有的教科書採用這一系統。 系統演化的兩大學派關於最古老被子植物的形態特徵問題,原始類群和進化類群各自具有的形態特徵問題,長期以來成為植物分類學家研究的中心,爭論的焦點,尤其是對被子植物的“花”的來源上,意見分歧最大,形成兩個學派,即所謂“假花學派”和“真花學派”,或者稱“柔荑學派”和“毛茛學派”。前者以德國植物學家恩格勒為代表,後者以美國植物學家柏施及哈利爾,英國植物學家哈欽松為代表。
認為被子植物的花和裸子植物的球花完全一致,每個雄蕊和心皮,分別相當於1個極端退化的雄花和雌花,因而構想被子植物是來自棵子植物麻黃類的彎柄黃Ephedra campylopoda,如下圖所示,在這個構想里,雄花的苞片變為花被,雌花的苞片變為心皮,每個雄花的小苞片消失後,只剩下一個雄蕊,雌花小苞片消失後只剩下胚珠,著生於子房基部。由於裸子植物,尤其是麻黃和買麻藤等都是以單性花為主,因而構想原始被子植物是具單性花。這種理論稱為假花學說(Pseudanthium theory)。根據此說,現代被子植物的原始類群,應該具有單性花、無被花、風媒花和木本的柔夷花序類植物,如木麻黃目、胡椒目、楊柳目等。這一學派的觀點為現代多數系統學家所反對,根據解剖學、孢粉學等研究資料證明柔荑花序類應為次生類群。
相關詞條
植物學
中國被子植物中含100種以上的科
| 數量 | 名稱 | 拉丁學名 |
| >2000 | 菊科 | Compositae |
| 1500~2000 | 豆科 | Leguminosae |
| 1000~1500 | 禾本科 | Grmineae |
| 薔薇科 | Rosaceae | |
| 蘭科 | Orchidaceae | |
| 800~1000 | 唇形科 | Labiatae |
| 毛茛科 | Ranunculaceae | |
| 500~800 | 莎草科 | Cyperaceae |
| 杜鵑花科 | Ericaceae | |
| 玄參科 | Scrophulariaceae | |
| 傘形科 | Umbelliferae | |
| 400~500 | 十字花科 | Cruciferae |
| 大戟科 | Euphorbiaceae | |
| 龍膽科 | Gentianaceae | |
| 苦苣苔科 | Gesneriaceae | |
| 樟科 | Lauraceae | |
| 報春花科 | Primulaceae | |
| 茜草科 | Rubiaceae | |
| 虎耳草科 | Saxifragaceae | |
| 300~400 | 紫草科 | Boraginaceae |
| 石竹科 | Caryophyllaceae | |
| 百合科 | Liliaceae | |
| 楊柳科 | Salicaceae | |
| 山茶科 | Theaceae | |
| 200~300 | 天南星科 | Araceae |
| 蘿藦科 | Asclepiadacaeae | |
| 小檗科 | Berberidaceae | |
| 忍冬科 | Caprifoliaceae | |
| 景天科 | Crassulaceae | |
| 殼斗科 | Fagaceae | |
| 紫堇科 | Fumariaceae | |
| 蓼科 | Polygonaceae | |
| 蕁麻科 | Urticaceae | |
| 100~200 | 爵床科 | Acanthaceae |
| 槭樹科 | Aceraceae | |
| 石蒜科 | Amaryllidaceae | |
| 番荔枝科 | Annonaceae | |
| 夾竹桃科 | Apocynaceae | |
| 冬青科 | Aquifoliaceae | |
| 五加科 | Araliaceae | |
| 鳳仙花科 | Balsaminaceae | |
| 秋海棠科 | Begoniaceae | |
| 桔梗科 | Campanulaceae | |
| 衛矛科 | Celastaceae | |
| 藜科 | Chenopodiaceae | |
| 鏇花科 | Convolvulaceae | |
| 葫蘆科 | Cucurbitaceae | |
| 木蘭科 | Magnoliaceae | |
| 野牡丹科 | Melastomataceae | |
| 桑科 | Moraceae | |
| 紫金牛 | Myrsinaceae | |
| 桃金孃科 | Myrtaceae | |
| 木樨科 | Oleaceae | |
| 鼠李科 | Rhamnaceae | |
| 芸香科 | Rutaceae | |
| 茄科 | Solanaceae | |
| 馬鞭草科 | Verbenaceae | |
| 堇菜科 | Violaceae | |
| 葡萄科 | Vitaceae | |
| 姜科 | Zingiberaceae |
