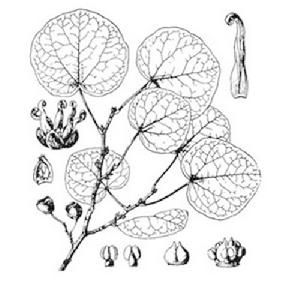
形態特徵
 長柄雙花木
長柄雙花木生長習性
 長柄雙花木
長柄雙花木分布範圍
 長柄雙花木
長柄雙花木種群生殖構件
 長柄雙花木
長柄雙花木種群數量動態
 長柄雙花木
長柄雙花木採用植物群落學的研究方法,對江西官山自然保護區長柄雙花木灌叢群落特徵及多樣性進行了研究。結果表明,群落結構特徵為單優勢種群落,喬木層種類少,樹冠不連續,灌木層以長柄雙花木占絕對優勢,林下更新較差。群落的物種多樣性指數低,表明生境條件特殊,群落結構不穩定。
優勢種群
運用種間聯結指數及2×2列聯表的X2統計量分別測定了長柄雙花木群落中物種總體關聯性和群落中主要樹種種對間的聯結性。結果表明:群落物種總體表現出一定程度的正關聯,群落處於比較穩定的階段。群落中13種主要樹種的78個種對中,有40個種對表現出正關聯,有37個種對表現出負關聯,1個種對表現為相互獨立的關係,正負關聯種對數比例接近1∶1。而長柄雙花木與其它12個物種之間有8對表現為正聯結,4對表現為負聯結,但均未達到顯著水平。根據種間聯結係數和群落結構,長柄雙花木分布群落中的13個主要優勢種分為3個生態種組。通過正交設計方法L9(34)研究了培養基中的植物生長調節劑6-BA、NAA、GA對長柄雙花木(Disanthuscercidifolavar.longipesH.T.Chang)不定芽生長速率的影響。實驗結果表明:最優水平組合為6-BA0.5mg·L-1 GA1.0mg·L-1.BA對實驗結果的影響達到極顯著水平(p≤0.01),該因素不同水平間差異達到極顯著水平(p<0.01)。
群落物種多樣性
分別運用Shannon-Wiener指數和物種均勻性指數探討了瀕危植物長柄雙花木群落的物種多樣性。研究結果表明,長柄雙花木群落物種多樣性水平隨海拔高度下降而呈上升趨勢。喬木層物種多樣性與群落穩定性有關,隨群落穩定性增加多樣性指數升高;而草、灌層物種多樣性則以喬木層蓋度高及群落生境優越者高。
繁育系統
 長柄雙花木
長柄雙花木花粉和資源限制
 長柄雙花木
長柄雙花木遺傳分化
 長柄雙花木
長柄雙花木生殖特性
 長柄雙花木
長柄雙花木開花物候指數與生殖間的相關分析表明,始花時間與開花數量、座果率及花期長度之間均具顯著負相關關係,而開花數量與花期長度之間則呈顯著正相關,但均為線性相關。長柄雙花木開花物候在種群間的差異和種群內年度間的相似性說明,其開花時間可能是由與其相關的複雜的微生境特徵和(或)由其遺傳因子決定的,同時也反映了種群間的遺傳分異和種群內個體間的遺傳一致性。作為一種瀕危物種,長柄雙花木在這種環境的選擇壓力之下,形成了“大量、集中開放的花”的開花模式,吸引到更多的傳粉者的訪問,從而達到生殖成功。
光合功能
 長柄雙花木
長柄雙花木研究了生長在不同光條件下的長柄雙花木幼苗葉片含水量、質膜相對透性、光合色素含量及光合速率的變化。結果表明,組織含水量、自由水含量及滲透勢均以3層遮蔭處理最高,並有隨遮蔭程度的減少而下降的趨勢。幼苗的淨光合速率在晴天表現為雙峰型日變化特徵,且在自然光照下的幼苗“午休”現象更為明顯。分析表明,長柄雙花木幼苗為喜蔭類型,而其分布地人為形成的裸露地不能為其提供足夠的蔭蔽環境,致使居群更新能力不強,這可能是導致長柄雙花木分布地逐漸縮小而瀕危的原因之一。
光強的適應
在光強較高的純林和闊葉林內,長柄雙花木淨光合速率呈“雙峰曲線”,並伴隨有胞間CO2的下降和氣孔限制值的增大;隨著日間光強的增大,葉片PQ還原程度和非光化學猝滅(NPQ)增加,φPSⅡ則下降,但後者午後隨著光強的下降可得到完全恢復;葉片吸收光能分配於光化學反應的比例隨著光強的增大而迅速減少;而天線色素熱耗散部分及反應中心過剩的光能則顯著增加,在午後光強減弱後兩者均可恢復到早晨的初始水平。在低照度光的竹林內,淨光合速率呈現“單峰曲線”,葉片PQ還原程度、NPQ以及φPSⅡ均無明顯變化,葉片吸收光能也主要用於光化學反應,熱耗散及過剩光能比例均較低。
瀕危及保護
 長柄雙花木
長柄雙花木雙花木系孓遺的單種屬植物。本種原變種產於日本,本變種為中國-日本植物區系的替代種,對探索植物系統發育和東亞植物地理方面具有一定的科學意義。雙花木屬僅雙花木一種,產於日本南部山區,長柄雙花木是它的變種,產於中國南嶺山地。目前,由於產地森林的砍伐破壞,長柄雙花木不僅個體數量越來越少,而且適於其生存的區域也日漸狹窄,已成為瀕危物種。
保護措施
湖南宜章莽山已建立自然保護區,應加強對本種的管護。浙江龍泉鳳陽山自然保護區和杭州植物園已引種繁殖與遷地保存。
栽培方法
本種尚無栽培記錄。可用種子育苗,或試用嫩枝扦插。人工繁殖長柄雙花木,以彎枝壓條和空中壓條成活率最高,達95%以上。
常見病蟲害
 長柄雙花木葉枯病
長柄雙花木葉枯病綜合管理:
(1)合理密植,合理施肥,增強生長勢。
(2)零星發生不防治,但要及時清除病殘體。
(3)連片侵染髮病時,用50%硫懸浮劑200-400倍液,27%銅高尚500-800倍液,3%多抗霉素900倍液,25%阿密西達懸浮劑1250-2500倍液噴霧。
 長柄雙花木炭疽病
長柄雙花木炭疽病綜合管理:
(1)增施有機肥,北方防寒;南方雨水多及時排水,防止濕氣滯留,適量增施磷鉀肥。
(2)及時修剪病枝葉和清除病殘體,集中深埋或燒毀。
(3)零星發生不防治,連片侵染髮病時,用29%石硫合劑100-200倍液,30%懸浮劑鹼式硫酸銅400-500倍液(發病前施藥),27%銅高尚500-800倍液,4%農抗120水劑600-800倍液,25%阿密西達懸浮劑100-200mg/L濃度藥液,20%噻菌銅500-800倍液噴霧。
 長柄雙花木褐斑病
長柄雙花木褐斑病綜合管理:
(1)及時清除病殘體,集中深埋或燒毀。
(2)合理密植,適量增施磷鉀肥。
(3)連片侵染髮病時,用3%多抗黴素900倍液,29%石硫合劑100-200倍液,27%銅高尚500-800倍液,4%農抗120水劑600-800倍液,1%武夷菌素100-200倍液噴霧。
植物進化史
(一)7月份葉芽開始分化,9月初葉芽長0.6厘米,10月底至11月初葉芽生長最快,此後停止生長,進入休眠期。第二年3月中旬葉芽恢復生長,下旬生長最快,4月上旬葉芽開放展葉,4月中旬全部展葉完畢。9月下旬葉片轉變為紅色,11月初開始落葉,元月初葉子落完。長柄雙花木葉片的生長節律十分奇特,一片葉子在展葉後的兩天內完成葉面積63%的增長,生長速度最快;10天后葉片的長和寬基本趨於穩定,不再生長。
(二)5月初,花芽開始分化;7月中旬花芽長0.2厘米;9月上旬花芽生長較快,9月下旬花芽膨大開花;10月中旬花瓣開始凋謝,10月底落花。因此,幼果是在冬天度過的,飽滿的種子少,空粒多,致使天然林下較少發現幼苗。這種現象不利於長柄雙花木的繁衍,也是導致其瀕危的原因之一。在冬天,幼果為了抵抗惡劣氣候的打擊,必須積累大量的疏水性物質而形成堅硬的種皮,儘管這種堅硬的外殼不利於以後的出苗,但比“全部凍死”要好。幼果在第二年6-7月生長最快,9月下旬轉黃色,10月上旬成熟。
金縷梅目百科(一)
| 金縷梅目(Hamamelidales) 雙子葉植物綱(木蘭綱) 木本。單葉互生,稀對生,多有托葉。花兩性、單性何株或異株,排成總狀花序、頭狀花序或柔荑花序;異被、單被或無被;雄蕊多數至定數;子房上位至下位,心皮1至多數,離生或合生. |
國家二級保護植物
| 其他 | 蟲草、鬆口蘑、髮菜 |
| 蕨類植物 | 鹿角蕨、桫欏科、亨利原始觀音座蓮 |
| 被子植物 | 鵝掌楸、香木蓮、降香黃檀、閩楠、長柄雙花木、長喙厚朴、野大豆、連香樹、西康玉蘭、藥用野生稻、舟山新木姜子、胡豆蓮、翅果油樹、羊角槭、絨毛皂莢、鹽樺、瓣鱗花、版納黑檀、浙江楠、油丹、永瓣藤、格木、普陀樟、普通野生稻、無翼坡壘、山銅材、寶華玉蘭、大果木蓮、大葉木蓮、大葉木蘭、地楓皮、圓葉玉蘭、四數木、台灣水青岡、厚朴、華南錐、半楓荷、十齒花、凹葉厚朴、任木、東京桐、三棱櫟、七子花 |
| 裸子植物 | 黃杉屬、金錢松、翠柏、紅檜、篦子三尖杉、秦嶺冷杉、福建柏、海南油杉、毛枝五針松、朝鮮崖柏、岷江柏木、太白紅杉、大果青扦、大別山五針松、四川紅杉、華南五針松 |
