
概述
 真菌界
真菌界生物的1界。本界成員均屬真核生物,它是真菌的最高分類階元。真菌廣泛分布於全球各帶的土壤、水體、動植物及其殘骸和空氣中,營腐生、寄生和共生生活。自C.von林奈把生物區分為動物和植物兩界以後的200多年以來,真菌因固著生活和細胞具壁而一直被歸入植物界。從20世紀中葉起,生物學家認為真菌的起源、組織、營養方式和細胞壁的組分等都與植物不同,把它歸入植物界並不妥當。真菌營養方式是“吸收異養型”,主要作用是分解,它與植物的光合自養和動物的攝食異養,都有著本質的區別。所以,在近20~30年的4界以上分類系統中,大多將真菌獨立成界。
真菌種屬很多,已報導的屬達1萬以上,種超過10萬個。其營養體除少數低等類型為單細胞外,大多是由纖細管狀菌絲構成的菌絲體。低等真菌的菌絲無隔膜,高等真菌的菌絲都有隔膜,前者稱為無隔菌絲,後者稱有隔菌絲。在多數真菌的細胞壁中最具特徵性的是含有甲殼質,其次是纖維素。常見的真菌細胞器有:細胞核、線粒體、微體、核糖體、液泡、溶酶體、泡囊、內質網、微管、鞭毛等;常見的內含物有肝糖、晶體、脂體等。
真菌的營養菌絲常發生多種變態,從而更有效地獲取養料,以滿足生長發育的需要。常見的變態菌絲有吸器、壓力胞、菌網和菌套等。前兩者多在植物寄生菌中形成,後兩者多在捕食線蟲等的真菌中出現。有些真菌在發育的某個階段,菌絲體變成比較疏鬆的或緊密交織的組織,這與通常組成菌體的疏鬆菌絲不同,叫做密絲組織。這種組織構成許多真菌不同類型的結構,如菌索、菌核和子座等。
真菌在生活中所需要的有機物質都依賴於自然界的其他生物。從死有機體中吸取養料的真菌叫做腐生菌。能侵害活的有機體、而不能生活在死有機體上的真菌叫做絕對寄生菌。寄生和腐生並不是絕對的,在一定條件下,一些真菌既能侵害活有機體又能生活在死有機體上,這種真菌叫做兼性寄生菌或兼性腐生菌。有許多真菌一方面從其他活有機體攝取養料,一方面又向同一活有機體提供養料或好處,這是一種共生現象,具有共生關係的真菌稱共生菌。
真菌的繁殖方式包括無性生殖、有性生殖和準性生殖。無性生殖是指不經過兩性細胞的結合便產生新的個體。生殖方法有:①體細胞(菌絲)的斷裂;②體細胞分裂成子細胞;③體細胞或孢子的芽殖;④各種無性孢子(如遊動孢子、孢囊孢子、分生孢子、厚垣孢子等)的產生,每個孢子可萌生芽管,再形成菌絲體。無性生殖在真菌的繁衍和傳播上起重要作用。有性生殖有遊動配子配合、配子囊接觸交配、配子囊交配、性孢子配合和體細胞配合等形式。有的真菌在質配後立即進行核配。另一些真菌不是立即進行核配,因此出現一個雙核階段,即每個細胞里有兩個沒有結合的核,細胞生長或分裂時,它們同時分裂,這是真菌特有的現象。這兩個核一直要到相當晚的時候才能配合。核配後或遲或早會發生減數分裂,重新使染色體的數目減為單倍。真菌的有性生殖最後形成各種有性孢子,如休眠孢子囊、卵孢子、接合孢子、子囊孢子和擔孢子。
1952年,美國的C.蓬泰科爾沃和羅珀在絲狀真菌中發現的一種導致基因重組的機制。在這種機制中,遺傳性的重組不是依賴有性生殖的減數分裂,而是依賴準性生殖的有絲分裂。準性生殖的過程可概括為質配、核配和單倍體化幾個階段。準性生殖在演化中具有一定的重要性。有性生殖和準性生殖並不互相排斥,有的真菌,如構巢麴黴,既發生有性生殖也發生準性生殖。有的生活在水域,有的生活在陸地;有的生長在熱帶,有的生長在寒帶;有的全年都能生長,有的只在某個季節才生長出來;有的寄生於其他生物,有的與其他生物共生,也有的只能生長在其他生物的腐敗殘骸上。每種真菌對養料、溫度、濕度、酸鹼度、氧、光線等都有特殊的要求。
關於真菌的起源和演化主要有兩派看法。一派認為真菌是由藻類演化而來。這些藻類因喪失色素而從自養變成異養,生理的變化引起了形態的改變。另一派認為除卵菌來自藻類外,其餘的真菌來自原始鞭毛生物。
研究歷史
 真菌界
真菌界真菌是一個古老的生物類群,在泥盆紀(4億多年前)的第一批化石植物中,就有腐生和寄生的真菌。中國古代文獻記載有黃帝與歧伯討論醪醴(即酒)、夏代儀狄釀酒、周代杜康制酒的傳說,從龍山文化出土的陶器中有盛酒的樽、沖酒的盉、煮酒的斝,這表明中國在新石器時期已經有了酒。宋代陳仁玉所著《菌譜》(1250)描述了食用菌11種,明代潘之恆所著《廣菌譜》(1500)描述了食用菌19種。1977年在浙江餘姚縣河姆渡村進行考古發掘,出土物中就有菌類,這表明在仰韶文化時期(距今6000~7000年),中國人已經採食蘑菇。
在1700~1850年間,各國科學家廣泛開展對大型真菌的研究,義大利的P.A.米凱利、荷蘭的D.C.H.佩爾松、瑞典的E.M.弗里斯家族、英國的M.J.伯克利、捷克的C.J.柯爾達、法國的L.R.蒂拉納兄弟等,都對真菌學的發展有重大的貢獻。
1851~1950年間是近代真菌學全面發展的時期。德國植物學家H.A.德巴里由於對真菌的系統發育、多型性和生活史的精湛研究而被譽為近代真菌學的奠基人。義大利P.A.薩卡爾多的巨著《真菌彙刊》是這一時期對真菌分類學的巨大貢獻。
20世紀50年代以來,現代真菌學飛速發展,如:①研究真菌的細微結構;②套用新技術和新方法;③醫學真菌飛躍發展;④出現真菌毒素學;⑤抗癌真菌藥物受到重視;⑥創立分子真菌學;⑦發展食用真菌,提出健康食品的名稱;⑧改變生物分類法,將真菌獨立成界等。
20世紀國中國用現代方法研究真菌,1915年胡先驌發表《菌類鑑別法》,為中國真菌鑑定打下了基礎。戴芳瀾於1927年出版《江蘇真菌名錄》,於1930年發表南京釣絲殼新種,是中國人用現代方法研究中國真菌的第一個新種。1939年鄧叔群出版《中國高等真菌志》;1951年王雲章出版《中國鏽菌索引》。此外,還有不少植物學家、植物病理學家、工業真菌學家和醫學真菌學家,也從事一些有關真菌學的研究。1980年成立中國真菌學會,1982年創刊中國《真菌學報》,現已開始籌編中國真菌志。
意義:真菌是一項豐富的自然資源。人和動物每年消耗大量的真菌菌體和子實體(食用菌);真菌也是重要的藥材(藥用真菌)。
真菌的某些代謝產物在工業上具有廣泛用途,如乙醇、檸檬酸、甘油、酶製劑、甾醇、脂肪、塑膠、促生素、維生素等。而且這些東西都能進行大規模的生產(見有機酸發酵微生物、酶製劑微生物、維生素髮酵微生物、甾體化合物的微生物轉化作用)。
在農業、林業和畜牧業中,真菌既有有害的一面,又有有利的一面。真菌能引起植物多種病害,從而造成巨大的經濟損失。例如,1845年歐洲由於馬鈴薯晚疫病的流行摧毀了5/6的馬鈴薯,中國由於1950年的小麥鏽病和1974年的稻瘟病而使小麥和水稻各減產120億斤。但是,菌根能對多種木本和草本植物提供必不可少的營養。在真菌的腐解作用中,它使許多重要化學元素得以再循環。真菌直接或間接地影響著地球生物圈的物質循環和能量轉換。
結構特徵
 真菌界
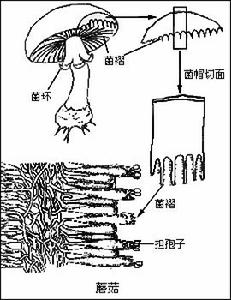
真菌界真菌是多型性的生物。它是以吸收水分和養料進行營養增殖的菌體,由簡單到複雜,一般可分為原質團、單細胞、假菌絲、兩型菌絲和菌絲體。菌絲體是常見的典型性營養體,由微小的絲狀物構成,它在基物上向一個方向分枝、延伸以便獲取養料。
1、細胞:菌絲細胞的四周包有透明的薄膜。細胞壁的裡面充滿原生質體以及浸沒在原生質體中的各種細胞器。細胞壁為多層薄片結構,主要化學成分是多糖,另外含有蛋白質、類脂以及其他物質。在多數真菌的細胞壁中最具特徵性的是甲殼質,其次是纖維素。細胞壁的內側是由蛋白質和類脂組成的連續結構質膜。位於質膜與細胞壁之間的須邊體,在真菌中也比在其他生物中更為常見。
在質膜包裹著的細胞質中,常見的真菌細胞器有:細胞核、線粒體 、微體 、核糖體、液泡 、溶酶體 、泡囊、內質網、微管、鞭毛等;常見的內含物有肝糖 、晶體、脂體等;沃羅寧體和脂體做超薄切片不易看見,而是實際上存在的一層薄膜。細胞核有核膜、核膜為雙層結構,並具有特徵性的核膜孔。細胞核常含有一個大部分為RNA所組成的核仁,當核分裂時,核仁有時消失。高爾基器在真菌中不經常出現,至少不以典型的形式出現。2、隔膜:大多數絲狀真菌的原生質是由橫隔膜隔開的,這種隔膜叫做分隔或隔膜。低等絲狀真菌的隔膜在繁殖器官的基部形成,繁茂的生長菌絲無隔多核或是無隔。如果菌絲變老時,隔膜可隨處形成。菌絲死亡時,原生質就移向生長點,隨即形成隔膜,將死的部分與活的部分分開,因此,真菌的原生質不死。
隔膜是由菌絲內壁向內生長而成,有的繼續生長,直至變成完整的隔片(封閉隔);有的發育不全,常留有一個孔(單孔隔)或多個孔(多孔隔),容許包括細胞核在內的一切細胞器通過,菌絲細胞變老或受傷時,旁邊的伏魯寧體和蛋白質晶體便可進行堵塞。多數擔子菌的隔膜有一中心孔,孔的邊緣膨大使中心孔成為琵琶桶狀的特殊結構,外面覆蓋一層弧形的膜,即為桶孔覆墊,覆墊有孔或管,由內質網形成。這種隔膜稱為桶孔隔膜。真菌隔膜的形態、結構、功能和性質涉及真菌核移機制和真菌細胞的基本概念,並可能有助於系統發育學說的發展。
3、菌絲的變態:在長期的自然選擇下,真菌的營養菌絲髮生多種變態,從而更有效地獲取養料,以滿足生長發育的需要。常見的變態菌絲有吸器、壓力胞、菌網和菌套等。前兩者多在植物寄生菌中形成,後兩者多在捕食線蟲等的真菌中出現。
4、組織:有些真菌在發育的某個階段,菌絲體變成比較疏鬆的或緊密交織的組織,這與通常組成菌體的疏鬆菌絲不同,叫做密絲組織。常見的密絲組織有長軸組織和擬薄壁組織兩種類型:前者頗為疏鬆,構成此種組織的菌絲大致平行排列,其典型的長形細胞容易識別;後者排列緊密,常是等徑或卵形,類似管束植物的薄壁細胞。這兩種組織構成許多真菌不同類型的營養結構,如菌索、菌核和子座等。
菌索是由菌絲體平行排列組成的長形索狀物,周圍有外皮,尖端為生長點,有幫助真菌運送物質和蔓延侵染的功能,而在不適環境下呈休眠狀態。菌核是堅硬的、能抵抗不良環境的休眠體,外壁由深色厚壁細胞構成,內層為淺色擬薄壁細胞。不同的真菌形成不同的菌核(如藥用的茯苓、豬苓、雷丸和麥角等)。它可以長期休眠,在適宜條件下重新萌發,產生子實體、菌絲和分生孢子。子座是堅實的營養結構,有時由菌絲和寄主組織混合構成,它很像座墊或床座,常常在座上或座中形成子實體。
營養與生長
 食用菌
食用菌真菌對營養的需要與動物和細菌相似,而不像綠色植物那樣能夠自己合成食物。
1、營養成分:真菌能產生各種水解酶,將糖類、澱粉、纖維素、木質素等碳水化合物以及蛋白質和脂肪分解,用作食物。大多數真菌能利用無機或有機氮以及各種礦物元素來合成自己的蛋白質。實驗研究表明:絕大多數真菌都需要碳、氫、氧、氮、磷、鉀、鎂、硫、硼、錳、銅、鉬、鐵和鋅,可能還需要鈣。許多真菌需要硫胺素和生物素,有些真菌需要吡哆醇和肌醇,但還沒有發現需要從食物中吸取核黃素的真菌。一般來說,葡萄糖是真菌最好的碳源,有機氮化合物是真菌最好的氮源。真菌的食物以植物性的物質為主,其次是動物性的物質。
2、營養方式:真菌在生活中所需要的有機物質都依賴於自然界的其他生物。從死有機體中吸取養料的真菌叫做腐生菌。這些養料一般稱為基物或基質。能侵害活有機體,而不能生活在死有機體上的真菌叫做絕對寄生菌。被寄生菌侵害的活有機體叫做寄主。近幾十年,人們已經開始設計能生長絕對寄生菌的合成培養基,例如一些鏽菌能在人工培養基上生長並完成它們的生活史。寄生和腐生並不是絕對的,在一定條件下,一些真菌既能侵害活有機體又能生活在死有機體上,這種真菌叫做兼性寄生菌或兼性腐生菌。有許多真菌一方面從其他活有機體攝取養料,一方面又向同一活有機體提供養料或好處,這是一種共生現象,具有共生關係的真菌叫做共生菌。
3、生長:主要指營養性菌體體積的加大和數量的增多。單細胞真菌菌體的生長,主要是經過細胞膨大、細胞核分裂、細胞質合成,最後達到細胞的芽殖或裂殖,進入無性繁殖。絲狀真菌的生長是以頂端延長的方式進行。菌絲頂部是菌絲體的生長點,長50~100微米,其中含有大量的泡囊、殼質體和細胞核等。泡囊以水泡狀形式從內質網轉移到高爾基器,在那裡濃縮、加工,並把它們的類內質網膜轉化為類質膜,然後從分散高爾基器分泌釋放,移至菌絲頂部,與質膜融合,使菌絲頂部的質膜增加。同時,頂部泡囊的內含物也被用來合成細胞壁,這些內含物含有壁的溶解酶類、壁的合成酶類、以及一些合成細胞壁的前體物。泡囊具有3種作用:①運輸細胞壁溶解酶去破壞原來壁組成之間的鍵;②運輸新的壁物質,在壁合成酶的作用下,將它併入細胞壁中;③在生長期間增加質膜的表面積。殼質體是比泡囊小得多的微泡囊結構,是運輸甲殼質合成酶的容器,為菌絲的伸長創造條件。在逐漸硬化的細胞壁和逐漸擴大的液泡的壓力下,菌絲中活躍的原生質從衰老部分流向頂端,使菌絲頂端不斷向前伸長。菌絲的這種生長方式叫做菌絲頂端生長。
繁殖形式
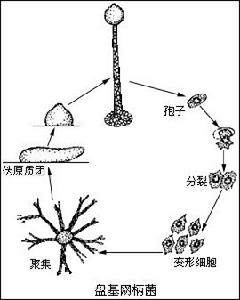
 真菌界
真菌界具有種的典型特徵的新個體的形成。真菌的繁殖器官由營養體產生或轉變而來。有些真菌在形成繁殖器官時,整個菌體轉變成一個至多個繁殖結構,這叫做整體產果式,這種結構分化少,比較原始。大多數真菌的繁殖器官只是由菌體的一部分構成,其餘部分繼續維持正常的營養活動,這叫做分體造果式。真菌的繁殖包括無性生殖、有性生殖和準性生殖。
1、無性生殖:菌的無性生殖是指不經過兩性細胞的結合便產生新的個體。生殖方法有:①體細胞(菌絲)的斷裂;②體細胞分裂成子細胞;③體細胞或孢子的芽殖;④各種無性孢子(如遊動孢子、孢囊孢子、分生孢子、厚垣孢子等)的產生,每個孢子可萌生芽管,再形成菌絲體。無性生殖在真菌的繁衍和傳播上起重要作用。
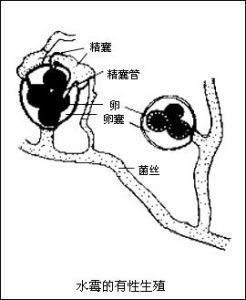
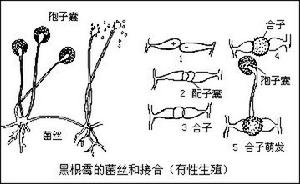
2、有性生殖:菌的兩個可親和性細胞核的結合。這種核的結合是通過遊動配子配合、配子囊接觸交配、配子囊交配、性孢子配合和體細胞配合來實現的。典型的有性生殖過程包括3個明顯不同的階段:第一階段叫質配,即兩個帶核的原生質體在同一細胞內互相配合;第二階段叫核配,即上述細胞里兩核的配合。有的真菌在質配後立即進行核配。另一些真菌不是立即進行核配,因此出現一個雙核階段,即每個細胞里有兩個沒有結合的核,細胞生長或分裂時,它們同時分裂,這是真菌特有的現象。這兩個核一直要到相當晚的時候才能配合。核配後或遲或早會發生減數分裂,重新使染色體的數目減為單倍,這就是有性生殖的第三階段。
真菌的有性生殖最後形成各種有性孢子,如休眠孢子囊、卵孢子、接合孢子、子囊孢子和擔孢子。這些孢子是許多真菌用以渡過不良環境的休眠體,也是許多植物病原真菌每次初侵染的傳染體,對於真菌的復壯或形成雜種優勢具有重要意義。同時,有性孢子也是真菌分類的重要依據。
根據有性生殖的親和性,真菌可分為同宗配合和異宗配合兩類:前者是每一菌體自身可孕,不靠其他菌體的幫助便可單獨進行有性生殖;後者是每一菌體自身不孕,不管是否雌雄同體,都需要藉助其他可親和性菌體的相對交配型來進行有性生殖。異宗配合又有兩極性和四極性之分。孤雌生殖在真菌中也時有發現。
3、準性生殖:1952年,美國的G.蓬泰科爾沃和羅珀在絲狀真菌中發現的一種導致基因重組的機制。在這種機制中,遺傳性的重組不是依賴有性生殖的減數分裂,而是依賴準性生殖的有絲分裂。準性生殖的第一步是形成異核體。異核體菌絲的形成有3種途徑:①異核菌絲的結合;②同質菌絲內的核發生突變;③菌絲內一些單倍體核進行核配形成二倍體核。準性生殖的第二步是兩個核的融合,同質的和異質的核都可以核配,形成同質的二倍體核和異質的二倍體核,結果,這個階段的菌絲細胞中最少含有5類核:兩種單倍體核,兩種純合二倍體核,一種異合二倍體核。但是,自發形成異合二倍體核的頻率非常低,如構巢麴黴在100萬個菌株中才出現一個異合二倍體。準性生殖的第三步包括二倍體核一系列非典型的、不規則的單倍體化。由於有絲分裂後期姊妹染色單體未分開,或者由於一個染色體分裂為2以後都趨向一極,而造成子核的染色體數目不相等,產生非整倍體,第1個子核多一個染色體(2n+1),第2個子核又少一個染色體(2n-1)。“2n+1”非整倍體又稱三體,往往失去一個染色體而成為二倍體。在這個過程中,可以由雜合體變為純合體。“2n-1”非整倍體又稱單體,常在不斷的有絲分裂中繼續喪失染色體,直至成為單倍體,準性生殖全過程可概括為質配、核配和單倍體化。半知菌不發生有性生殖,發生準性生殖的也只有麴黴、青黴和鐮孢霉屬中的少數種。準性生殖在演化中具有一定的重要性。有性生殖和準性生殖並不互相排斥,有的真菌,如構巢麴黴,既發生有性生殖也發生準性生殖。
 真菌界
真菌界在異合二倍體繁殖的有絲分裂中,染色體之間偶爾也發生交換,形成新的組合和連鎖(見連鎖和交換)。有些新組合的二倍體核在形成分生孢子時進入孢子,而在孢子萌發時生出二倍體菌絲。這樣的重組是準性生殖的主要方面,因為它使沒有有性生殖的真菌也能得到有性生殖的益處。不過發生的頻率很低,僅為有性生殖的1/500。
準性循環的遺傳雖然具有有性生殖的內容,但與有性循環之間還有幾點區別:①準性循環的過程類似有性循環,但不像有性循環那樣固定,而且單倍體化的完成要犧牲大量的非整倍體;②在一個菌絲細胞內,異核現象和異合現象可以同時發生,不過異合機會極少,必須進一步分離才能發現。由於一個二倍體自發分離(體細胞交換和單倍體化)的頻率很低,往往需要用各種化學的或物理的因子加以誘導,例如用對氟苯丙氨酸(PFA)誘導構巢麴黴單倍體化就有特別顯著的效果,用氟尿嘧啶(FU)誘導構巢麴黴的有絲分裂交換(體細胞交換),效果也很明顯。另外還有許多因子,如氮芥、甲醛和紫外線等,對構巢麴黴的單倍體化和有絲分裂交換都起誘導作用。
準性生殖的發現進一步深化了對真菌的認識。通過體細胞重組,首先可以在無性生殖的真菌中進行基因在染色體上的定性研究,其次可以在半知菌中進行雜交育種。在半知菌的演化中,有些似已固定用準性生殖來代替有性生殖了。
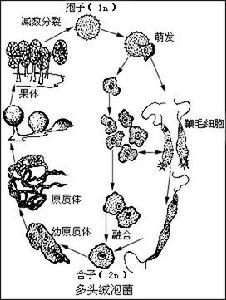
4、核循環:像其他生物一樣,真菌通常也有一個相當於綠色有機體配子體和孢子體的單倍體和二倍體結構的循環(圖2)。二倍體階段由核配開始,到減數分裂完成。但是,在大多數真菌中,菌體並沒有明顯的單倍體和二倍體的交替,代表二倍體階段的僅僅是合子。
生態習性
 真菌界
真菌界天然真菌或人工培養真菌與生存條件之間的相互關係。真菌種類很多,對生存條件的要求和反應各不相同。它們有的生活在水域,有的生活在陸地;有的生長在熱帶,有的生長在寒帶;有的全年都能生長,有的只在某個季節才生長出來;有的寄生於其他生物,有的與其他生物共生,也有的只能生長在其他生物的腐敗殘骸上。這些不同的生活方式和生長習性,都是在漫長的發育歷史中形成的。每種真菌對養料、溫度、濕度、酸鹼度、氧、光線等都有特殊的要求。
1、腐生真菌及其習性:腐生真菌是數量多、分布廣、作用大的一群真菌,能把禁錮在有機體內的化學元素歸還給大自然,使碳素循環順利進行。當它們的菌絲與基物緊密接觸時,在外酶的作用下,可將周圍不溶性的有機物水解為可溶性的有機物,將大分子的有機物分解為小分子有機物,並通過菌絲壁的直接擴散作用進行吸收。首先吸收的是可溶性的糖和胺基酸。大多數真菌能夠迅速分解簡單的碳水化合物,如糖類、澱粉、半纖維素和某些蛋白質。對纖維素、脂肪和油類的分解比較慢,而且常常需要特殊的真菌類型。木質素、蠟和單寧最抗分解,只有少數真菌才能分解它們。
2、土壤真菌:土壤中有豐富的腐生真菌基質。真菌主要分布於距地表24厘米以內的土壤中,在0~10厘米間最多。它們的數量和種類隨土層的加深而遞減。
3、水生真菌:水體中生活並能完成生活史的真菌。
4、嗜熱真菌:40℃以上能生長繁殖的真菌。常見的有毛霉、毛殼菌、籃狀菌、嗜熱子囊菌等屬的一些種。農業上利用它們製造堆肥和栽培蘑菇時製備培養基。
5、喜冷真菌:10℃以下能生長的真菌。它們生長在極地、高山、雪下和冷藏庫中。南極的月平均溫度一般在0℃以下,但真菌比較普遍,已發現有結實串孢霉、總絲霉等多屬半知菌和酵母。在海拔4000米的高山和雪線附近也有蘑菇子實體生長。冷藏庫的溫度一般都在0℃以下,但在貯藏的肉類和果蔬上常有枝孢、枝霉、青黴和毛霉等真菌為害。
6、大氣真菌:空氣不像土壤和水體那樣能作為真菌的生活環境,但源出陸地的某些真菌菌絲的碎片和孢子卻構成了空中孢子區系。大氣真菌主要指對流層及其以上10公里處的真菌,只有少數的同溫層中。空中真菌的類型和密度,因季節、時間、氣候和人類的活動而發生變化。一般來說,在乾旱氣候的白晝,枝孢和鏈格孢等乾孢子類真菌占優勢;在潮濕的夜間,子囊孢子和擔孢子比較多。
7、寄生真菌及其寄主:植物寄生真菌:幾乎所有的植物都受真菌的侵襲和危害。受害最大的是栽培植物,往往造成減產和品質降低。
8、動物和人的寄生真菌:真菌能侵染各種動物和人,引起病害。使高等動物受害的絕大多數為半知菌,只有少數是接合菌和子囊菌。這些病原真菌都是兼性寄生菌,大多可以從土壤中分離出來。從人體分離出來的真菌約120種,其中半數是常見的腐生菌。至今還沒有發現專性寄生於人的真菌。
9、真菌寄生真菌:從另一種真菌獲取養料的真菌。已知有30多種壺菌寄生於其他壺菌和卵菌上。有些毛霉目的真菌常寄生於毛霉目的其他成員上,並具有高度的專化性。有些半知菌是卵菌、接合菌、子囊菌、擔子菌和其他半知菌的寄生物。這類真菌可用於害菌的生物防治。
10、共生真菌及其夥伴:同其他生物生活在一起,彼此受益的真菌(如地衣、菌根、蟲道真菌)。
起源與演化
真菌的起源和演化眾說紛紜,但直到現在還沒有一個能被真菌學家們普遍接受的學說。
一個學派認為真菌是由藻類演化而來。這些藻類因喪失色素而從自養變成異養,生理的變化引起了形態的改變。他們根據性器官的形態和交配方式進行推測,認為鞭毛菌來自綠藻,接合菌來自接合藻,子囊菌來自紅藻,擔子菌來自子囊菌。
另一個學派認為除卵菌來自藻類外,其餘的真菌來自原始鞭毛生物。鞭毛生物水生,具有一至數根鞭毛,有的含葉綠素或紅色素,有的含其他色素或無色素。含葉綠素的演化為藻,無色素的演化為菌。
20世紀中葉以前,有關真菌系統發育的構想、方案和系統樹,除生態習性和營養方式外,主要是以形態學為基礎。現在人們普遍認識到多數活有機體的基礎生物化學是相同的,而其差異如細胞壁的組分、DNA和RNA的含量、DNA中的G+C克分子百分含量、胺基酸的排列順序、血清反應以及某些代謝產物的含量與合成途徑等,則有利於確定它們之間的親緣關係。例如,根據血清反應,推測藻狀菌是管藻目中靠近無隔藻的兩個早期分支,後來又衍生出許多支系,直至今天的炭角菌、羊肚菌、鏽菌、鳥巢菌、牛肝菌,鵝膏菌等類群。再如,根據賴氨酸的生物合成途徑,推測壺菌可能是從與眼蟲藻有關的鞭毛生物演化而來。壺菌喪失鞭毛以後,由於菌絲髮達而演化成接合菌,
從接合菌再演化為子囊菌和擔子菌。所以,真菌的演化途徑可能是:與眼蟲藻有關的鞭毛生物→壺菌→接合菌→子囊菌→擔子菌。這是真菌演化的主軸,相當於綠色植物中的:綠藻→輪藻→苔蘚→維管束植物。過去認為卵菌是在產生高等真菌的演化線上的一個早期分支,現在卻認為它屬於另一個演化線,大概起源於與金藻有關的無色素類型,因而被排除在真菌以外。
下屬分類
真菌界以下的分類系統,有的分亞界,有的分門,有的直接分為綱,因作者而異。其中,英國真菌學家G.C.安斯沃思等人(1973)的分類系統,將真菌界分為2門、5亞門、20綱,既照顧了傳統習慣,又反映了發展趨勢,比較可取。
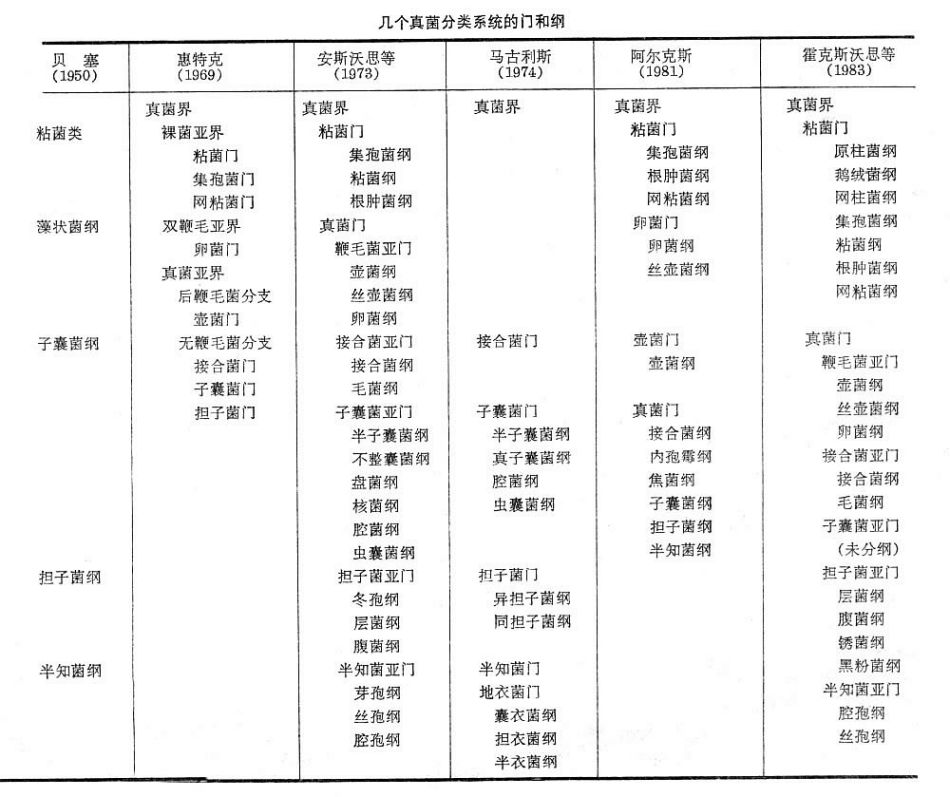 幾個真菌分類系統的門和綱
幾個真菌分類系統的門和綱真菌界以前的分類學家把他們隸屬於植物界,1973年後有的分類學家單分為一界,這類生物不能像植物一樣含有葉綠素,不能自制養料,又不能像動物一樣活動攝取食物,基本是從其他生物、生物屍體或排泄物上攝取營養,營寄生或腐生生活。主要分為兩門:粘菌門和真菌門。有的分類學家將真菌列為植物類的一門,既真菌門。
真菌界也可分為2個亞界,即粘菌亞界和真菌亞界:
1、粘菌亞界
粘菌生長在陰濕土壤、木塊、腐朽植物體、糞便等上面,細胞沒有壁,單核或多核。原生動物學家根據粘菌有變形蟲樣的單細胞階段,並能吞食固體顆粒,主張把粘菌放入原生動物之中。但是粘菌有多細胞階段,它們除吞噬營養外,也能吸收有機物,所以真菌學家也歡迎它們。已知粘菌約有500餘種。
2、真菌亞界:真菌和細菌一樣,也是自然界中強大的有機物分解者。它們以動、植物屍體、枯木爛葉為食物源,也可侵入活的生物體內攝取營養。有些真菌可和藻類等他種生物組成互利的結合體,如地衣、菌根等。除酵母菌等少數單細胞真菌外,大多真菌是多細胞的。這些真菌形態上的一個共同特徵是菌絲。真菌的身體其實就是分支或不分支的菌絲或由菌絲構成的菌絲體。菌絲是特殊形式的細胞,可長可短,其中有細胞核和細胞質。有些真菌的菌絲中有橫隔,將菌絲隔成一系列細胞,每細胞中或有一核或有二核,隨不同真菌而不同。有些真菌的菌絲中無橫隔,菌絲成為一個多核細胞。多數真菌細胞壁的主要成分是幾丁質。
