
 香石竹環斑病毒
香石竹環斑病毒基本資料
學名 Carnation ringspot virus
異名 Anjermozaiek virus,
英文名 Carnation ringspot virus
分類地位 番茄叢矮病毒科香石竹環斑病毒屬 Tombusviridae Diathovirus
分布
一般香石竹生長區CRSV都有分布。歐洲東部和中部的葡萄、堅果已報導有CRSV。歐洲:丹麥、芬蘭、法國、德國、義大利、立陶宛、荷蘭、波蘭和英國有分布。西半球:巴西、加拿大、哥倫比亞、墨西哥和美國有分布。大洋州:澳大利亞和紐西蘭有分布。寄主植物
 香石竹環斑病毒
香石竹環斑病毒寄主範圍:
實驗條件下,CRSV可侵染25科共133種植物(Hollings & Stone,1970; Kleinhempel et al.,1980)。自然條件下,CRSV侵染香石竹,多數果樹(李樹、梨樹、蘋果、酸櫻桃和甜櫻樹),葡萄及果園裡的雜草,如繁縷。實驗寄主範圍較自然寄主廣(Hollings & Stone,1970)。較易機械傳播道草本植物,可系統侵染茄科、豆科、葫蘆科。非系統侵染的寄主更廣(Hollings & Stone,1970)。實驗條件下,CRSV引起系統侵染,寄主的接種葉產生以壞死斑為中心的環斑,系統葉產生黃花、壞死斑、壞死脈。
診斷寄主:
莧色藜(Chenopodium amaranticolor)、昆諾阿藜(C. quinoa)、美國石竹(Diathus barbatus)、千日紅(Gomphrena globosa)、菜豆 (Phaseolus vulgaris)、番杏 (Tetragonia expansa)、豇豆屬(Vigna unguiculata ssp. sinensis)。
繁殖寄主:
美國石竹(Diathus barbatus)和克利夫蘭煙(N. clevelandii)。
測定寄主:
莧色藜(Chenopodium amaranticolor)、昆諾阿藜(C. quinoa)、豇豆 (Vigna unguiculata ssp. sinensis)。
株系:
根據寄主不同反應來區別株系未見報導。報導的幾個株系是基於不同聚集特性。A株系由12個粒子聚集成一個對稱性二十面體,這12個粒子亞基又形成更高一級的聚集物。N株系和R株系各自形成兩個雙向類核聚集物(Tremaine,1976)。
危害情況
CRSV侵染香石竹引起環斑、斑駁、葉和花扭曲、畸變(Fig. 1, Hollings & Stone, 1970; Kowalska,1972; Lovisolo & Lisa, 1978)。當CRSV與香石竹斑駁病毒(Carnation mottle virus)共同侵染時,病症加重(Kemp,1964; Holling & Stone, 1965)。在侵染嚴重或易感香石竹品種中,可見葉尖壞死現象。一般來說,CRSV侵染不能使寄主植物致死,當溫度持續在15℃-20℃時,壞死或其它症狀則更嚴重。溫度低於20℃時,症狀稍弱。堅果和果園作物上的症狀較輕且很難發現(Hiruki,1987)。CRSV在梨樹上引起“Stong pit”(Richter,1978)。在酸櫻桃和蘋果上症狀稍弱(Kleinhempel,1980)。形態特徵
 香石竹環斑病毒
香石竹環斑病毒粒子結構:
CRSV粒子是直徑34納米的正二十面體,T=3(Fig. 8; Kalmakoff & Tremaine, 1967)。與番茄叢矮病毒屬和香石竹斑駁病毒屬相似,CRSV粒子具有不同的顆粒狀表面(Lommel,1995)。180個分子量為38kd的外科蛋白組成粒子外殼,包裹兩條分子量分別為3.8kb和1.4kb的基因組單鏈環狀RNA(Kendall & Lommel, 1992; Ryabov, 1994; Lommel, 1995)。
粒子組份
核酸:
ssRNA, 占病毒粒子重量20%。基因組由RNA-1和RNA-2兩條RNA分子組成(Fig. 9)。RNA分子如何包裝成病毒粒子還未知(Hamilton & Tremaine, 1996)。有些粒子可能只包含RNA-2。有些學者認為每個粒子包含1個RNA分子,而有的學者則認為存在兩種類型粒子,一種只包含單個RNA-1,而另一個包含三個RNA-2分子。RNA-1分子量為1.246′106,共3840核苷酸,各核苷酸組成為1037A(27.01%),903C(23.52%),972G(25.31%)和928U(24.17%)。RNA-1基因庫登錄號L18870(Ryabov, 1994)。RNA-2分子量為0.455106, 共1403各核苷酸,各核苷酸組成為380A(27.1%),364U(25.96%),318C(22.68%)和340G(24.25%),基因登錄號M88589(Kendall & Lommel, 1992)。
蛋白質:
病毒粒子開始膨脹後,外科蛋白就能分離(Tremaine & Dodds, 1985)。包含單條多態的病毒粒子,其蛋白質占顆粒重量的80%。多態分子量為37900,由345個胺基酸組成。
基因組特性
基於同屬RCNMV的研究,認為CRSV裸露的核酸具有侵染力。CRSV RNA-1在缺少RNA-2時可單獨在植物原生質體中進行複製(Paje-Manalo & Lommel,1980),但侵染植物需RNA-1和RNA-2的共同作用。RNA-1外科蛋白3’可由1.5kb亞基因組RNA體內表達(Fig.10;Zavriev, 1996)。三種dsRNA 聚集在感病細胞中,這三種dsRNA 分別是兩種基因組RNA和一種亞基因組RNA(Fig.9)。RNA-2上的一個序列元件可激活RNA-1合成亞基因組RNA(Sit,1998)。根據RCNMV推測CRSV RNA的5’有m7GpppA, 而3’無polyA尾巴。RNA-1和RNA-2上只有5’ 端2個核苷酸和離3’端20個核苷酸遠的26個核苷酸是相同的,且3’端相同的26個核苷酸形成類似莖環結構。
RNA-1編碼2個開讀框(ORFs)。第一個ORF(p88)開始於甲硫氨酸密碼子,離5’端58個核苷酸處(Fig. 10),p88ORF編碼一個769胺基酸的多肽,分子量為87776。P88ORF被類反轉錄病毒-1核糖體移碼元件中止(Kim & Lommel,1998),產生一個236胺基酸,分子量為27158的先多肽(p27)。P27於前一個88KD蛋白(236aa)蛋白一致(Kim & Lommel,1994)。RNA-1近3’端的ORF編碼一個345胺基酸的外科蛋白p38,分子量為37921。p38ORF後接由436個核苷酸組成的非編碼區。RNA-2隻含一個ORF,編碼303胺基酸,分子量為33730的移動蛋白(p34)(Kendall & Lommel, 1992)。ORF起始於第47個核苷酸,P34後接一個441核苷酸的非編碼區(Fig. 10)。在體內,RNA-1指導27kd、57 kd、88 kd和37 kd多肽的合成。57 kd多肽體內不可見(Kim & Lommei,1994)。RNA-1編碼的88 kd多肽含保守“甘氨酸-天冬氨酸-天冬氨酸(Gly-Asp-Asp)”GDD元件,GDD元件在依賴RNA聚合酶中均存在(Ryabov,1994)。27 kd和88 kd多肽的胺基酸序列具有相似性,番茄叢矮病毒科病毒都編碼複製酶。外科蛋白胞間移動不需外科蛋白參與,但病毒通過維管束形成系統侵染需參與。RNA-2編碼34 kd的移動,對病毒胞間和長距離運輸起作用。
生物學
 香石竹環斑病毒
香石竹環斑病毒血清學:
CRSV具有中強免疫原性,易獲得高質量的多抗血清(Hiruki,1987)。凝膠免疫擴散試驗測定效價高於1/1024。鑑定CRSV常用凝膠雙擴散(Kowalska,1972)和ELASA試驗(Lommel,1982)。病毒粒子注射兔子產生的抗體在凝膠擴散試驗中不與游離的蛋白亞基反應,而在間接ELASA中與其抗原反應(Tremaine & Dodds, 1985)。凝膠擴散試驗中可區分不同株系(Tremaine, 1976; 1984)。
CRSV是香石竹環斑病毒屬的典型株系,該屬還包括另外兩個病毒:紅三葉草壞死花葉病毒(red clover necrotic mosaic virus, RCNMV)和甜三葉草壞死花葉病毒(sweet clover necrotic mosaic virus, SCNMV)(Lommel, 1995)。CRSV與香石竹斑駁病毒(carnation mottle cirus, CarMV)易混淆,CarMV在經濟香石竹作物上廣為分布(Lommel, 1983a),且CarMV與CRSV易混合侵染(Kemp, 1964; Holling & Stone, 1965)。採用負染法電鏡觀察,CRSV與CarMV區分不開,但兩者的血清學有差異,且在千日紅(Gomphrena globosa)和香石竹(Dianthus barbatus)引發的症狀不同。CarMV是香石竹斑駁病毒屬的典型株系。香石竹義大利環斑病毒(carnation italian ringspot virus)為番茄叢矮病毒屬成員(Russo, 1987),但病毒名稱與CRSV類似。 與香石竹環斑病毒屬病毒相似,香石竹斑駁病毒屬病毒和番茄叢矮病毒屬病毒均屬於番茄叢矮病毒科病毒(Lommel, 1995)。
病毒汁液穩定性 :
美國石竹病汁液中,溫度高於60℃,CRSV感染率大為降低。經冰凍真空乾燥後的克力夫蘭煙病汁液,乾粉的毒力可保持6年。病毒致死溫度80℃(10分鐘),稀釋限點為10-5。20℃時,體外存活期為50-60天。室溫,真空條件下,CRSV在克力夫蘭煙(Nicotiana clevelandii)汁液中可存活6年以上(Holling & Stone, 1970)。
傳播途徑
介體傳播:CRSV不能由昆蟲和土壤真菌(Hiruki,1987)傳播。CRSV能否由線蟲傳,還未有定論。早期報導,有些線蟲種類如Longudirys elongatus長針線蟲(L. macrosoma)和劍線蟲(Xiphineme diversicandatum)能傳播CRSV(Fritzsche & Schmelzer,1967; Kegler,1977)。最近報導認為病毒粒子直接從受侵染植物根部釋放到土壤中,再從土壤傳到植物,不需昆蟲介體介入(Kegler,1981; Brown & Trudgill, 1984)。病毒粒子在感病果園裡的流水中可見(Koening, 1988, 1989)。CRSV可能通過根部的微傷口進入植物,所以植物根部線蟲和土壤菌類的寄主增加了土壤傳播的機率,但傳播並不僅僅依賴於線蟲和土壤菌類。自然界中,CRSV的傳播大多是由於植物間的接觸、土壤污染、農事操作等引起的(Lommel,1983b)。
檢疫與防治
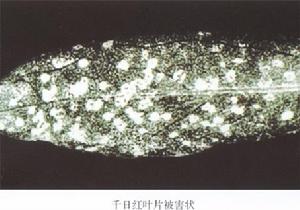 香石竹環斑病毒
香石竹環斑病毒檢驗方法
鑑別寄主反應 美國石竹(Dianthus barbatus L.)汁液摩擦接種,4~7天后接種葉表現局部壞死斑和環斑,接著為系統褪綠和半壞死,環斑。千日紅(Gomphrena globosa)。汁液摩擦接種,2~4天產生局部壞死斑,接著出現系統斑、斑駁和畸形。菜豆(Phaseolus vulgaris)汁液摩擦接種,4~5天后產生局部褪綠點,以後變為白色壞死、不規則系統斑點和壞死脈點,後長葉無症帶毒。莧色藜(Chenopodium amaranticolor)和昆諾阿藜(C.quinoa)汁液摩擦接種,2~4天后產生局壞死斑,通常無系統症狀。番杏(Tetragonia expansa)汁液摩擦接種,2~3天后產生局部白色壞死點,有時會發展為系統褪綠斑。豇豆(Vigna unguiculata ssp.sinensis)汁液摩擦接種,2~4天后產生局部壞死斑,隨著出現系統斑駁、壞死斑,葉片粗而捲曲。
血清學反應 凝膠擴散可檢出香石竹汁液的香石竹環斑病毒,雙抗體夾心 ELISA均為有效的檢測手段,凝膠擴散還可測出病毒株系的差異。ELISA可檢出1~10ng/ml的香石竹環斑病毒,間接ELISA較雙抗體夾心ELISA更靈敏。在理化性質方面,香石竹環斑病毒與三葉草壞死花葉病毒很相似,屬於一個病毒組,在間接ELISA試驗中兩者表現弱的血清學交叉反應,還有報導說香石竹環斑病毒與蕪菁黃花葉病毒組病毒有血清學交叉反應。
有關檢疫規定 香石竹環斑病毒是我國公布的《中華人民共和國進境植物檢疫危險性病、蟲、雜草名錄》中規定的二類危險性病毒,應嚴格限制引進繁殖材料數量,必須引進的應由出口國出具檢疫證書,進口後在指定的隔離溫室中檢查。
中國進境植物檢疫危險性病蟲草
| 時刻牢記外來入侵生物的危險性,知曉它們的習性,為減低危險做準備!!! |
