
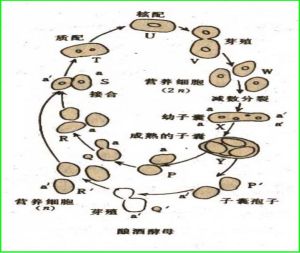
生活史
酵母的細胞有兩種生活形態,單倍體和二倍體。單倍體的生活史較簡單,通過有絲分裂繁殖。在環境壓力較大時通常則死亡。二倍體細胞(酵母的優勢形態)也通過簡單的有絲分裂繁殖,但在外界條件不佳時能夠進入減數分裂,生成一系列單倍體的孢子。單倍體可以交配,重新形成二倍體。酵母有兩種交配類型,稱作a和α,是一種原始的性別分化,因此很有研究價值。
基因組測序
 釀酒酵母
釀酒酵母釀酒酵母是第一個完成基因組測序的真核生物,測序工作於1996年完成。
釀酒酵母的基因組包含大約1200萬鹼基對,分成16組染色體,共有6275個基因,其中可能約有5800個真正具有功能。據估計其基因約有23%與人類同源。酵母基因組資料庫包含有酵母基因組的詳細注釋(annotation),是研究真核細胞遺傳學和生理學的重要工具。另一個重要的釀酒酵母資料庫由慕尼黑蛋白質序列信息中心維護。
在釀酒酵母測序計畫開始之前,人們通過傳統的遺傳學方法已確定了酵母中編碼RNA或蛋白質的大約2600個基因。通過對釀酒酵母的完整基因組測序,發現在12068kb的全基因組序列中有5885個編碼專一性蛋白質的開放閱讀框。這意味著在酵母基因組中平均每隔2kb就存在一個編碼蛋白質的基因,即整個基因組有72%的核苷酸順序由開放閱讀框組成。這說明酵母基因比其它高等真核生物基因排列緊密。如線上蟲基因組中,平均每隔6kb存在一個編碼蛋白質的基因;在人類基因組中,平均每隔30kb或更多的鹼基才能發現一個編碼蛋白質的基因。
酵母基因組的緊密性是因為基因間隔區較短與基因中內含子稀少。酵母基因組的開放閱讀框平均長度為1450bp即483個密碼子,最長的是位於XII號染色體上的一個功能未知的開放閱讀框(4910個密碼子),還有極少數的開放閱讀框長度超過1500個密碼子。在酵母基因組中,也有編碼短蛋白的基因,例如,編碼由40個胺基酸組成的細胞質膜蛋白脂質的PMP1基因。此外,酵母基因組中還包含:約140個編碼RNA的基因,排列在XII號染色體的長末端;40個編碼snRNA的基因,散布於16條染色體;屬於43個家族的275個tRNA基因也廣泛分布於基因組中。表1提供了酵母基因在各染色體上分布的大致情況。
基因組特徵
序列測定揭示了酵母基因組中大範圍的鹼基組成變化。多數酵母染色體由不同程度的、大範圍的GC豐富DNA序列和GC缺乏DNA序列鑲嵌組成。這種GC含量的變化與染色體的結構、基因的密度以及重組頻率有關。GC含量高的區域一般位於染色體臂的中部,這些區域的基因密度較高;GC含量低的區域一般靠近端粒和著絲粒,這些區域內基因數目較為貧乏。Simchen等證實,酵母的遺傳重組即雙鏈斷裂的相對發生率與染色體的GC豐富區相耦合,而且不同染色體的重組頻率有所差別,較小的Ⅰ、Ⅲ、Ⅳ和Ⅸ號染色體的重組頻率比整個基因組的平均重組頻率高。
 釀酒酵母
釀酒酵母酵母基因組另一個明顯的特徵是含有許多DNA重複序列,其中一部分為完全相同的DNA序列,如rDNA與CUP1基因、Ty因子及其衍生的單一LTR序列等。在開放閱讀框或者基因的間隔區包含大量的三核苷酸重複,引起了人們的高度重視。因為一部分人類遺傳疾病是由三核苷酸重複數目的變化所引起的。還有更多的DNA序列彼此間具有較高的同源性,這些DNA序列被稱為遺傳豐余(genetic redundancy)。
酵母多條染色體末端具有長度超過幾十個kb的高度同源區,它們是遺傳豐余的主要區域,這些區域至今仍然在發生著頻繁的DNA重組過程。遺傳豐余的另一種形式是單個基因重複,其中以分散類型最為典型,另外還有一種較為少見的類型是成簇分布的基因家族。成簇同源區(cluster homology region,簡稱CHR)是酵母基因組測序揭示的一些位於多條染色體的同源大片段,各片段含有相互對應的多個同源基因,它們的排列順序與轉錄方向十分保守,同時還可能存在小片段的插入或缺失。
這些特徵表明,成簇同源區是介於染色體大片段重複與完全分化之間的中間產物,因此是研究基因組進化的良好材料,被稱為基因重複的化石。染色體末端重複、單個基因重複與成簇同源區組成了酵母基因組遺傳豐余的大致結構。研究表明,遺傳豐余中的一組基因往往具有相同或相似的生理功能,因而它們中單個或少數幾個基因的突變並不能表現出可以辨別的表型,這對酵母基因的功能研究是很不利的。所以許多酵母遺傳學家認為,弄清遺傳豐余的真正本質和功能意義,以及發展與此有關的實驗方法,是揭示酵母基因組全部基因功能的主要困難和中心問題。
基因研究
 釀酒酵母
釀酒酵母因為釀酒酵母與同為真核生物的動物和植物細胞具有很多相同的結構,又容易培養,酵母被用作研究真核生物的模式生物,也是目前被人們了解最多的生物之一。在人體中重要的蛋白質很多都是在酵母中先被發現其同源物的,其中包括有關細胞周期的蛋白、信號蛋白和蛋白質加工酶。
釀酒酵母也是製作培養基中常用成分酵母提取物的主要原料。
酵母作為高等真核生物特別是人類基因組研究的模式生物,其最直接的作用體現在生物信息學領域。當人們發現了一個功能未知的人類新基因時,可以迅速地到任何一個酵母基因組資料庫中檢索與之同源的功能已知的酵母基因,並獲得其功能方面的相關信息,從而加快對該人類基因的功能研究。研究發現,有許多涉及遺傳性疾病的基因均與酵母基因具有很高的同源性,研究這些基因編碼的蛋白質的生理功能以及它們與其它蛋白質之間的相互作用將有助於加深對這些遺傳性疾病的了解。此外,人類許多重要的疾病,如早期糖尿病、小腸癌和心臟疾病,均是多基因遺傳性疾病,揭示涉及這些疾病的所有相關基因是一個困難而漫長的過程,酵母基因與人類多基因遺傳性疾病相關基因之間的相似性將為我們提高診斷和治療水平提供重要的幫助。
酵母作為模式生物的最好例子體現在那些通過連鎖分析、定位克隆然後測序驗證而獲得的人類遺傳性疾病相關基因的研究中,後者的核苷酸序列與酵母基因的同源性為其功能研究提供了極好的線索。例如,人類遺傳性非息肉性小腸癌相關基因與酵母的MLH1、MSH2基因,運動失調性毛細血管擴張症相關基因與酵母的TEL1基因,布盧姆氏綜合徵相關基因與酵母的SGS1基因,都有很高的同源性。遺傳性非息肉性小腸癌基因在腫瘤細胞中表現出核苷酸短重複順序不穩定的細胞表型,而在該人類基因被克隆以前,研究工作者在酵母中分離到具有相同表型的基因突變(msh2和mlh1突變)。
受這個結果啟發,人們推測小腸癌基因是MSH2和MLH1的同源基因,而它們在核苷酸序列上的同源性則進一步證實了這一推測。布盧姆氏綜合徵是一種臨床表現為性早熟的遺傳性疾病,病人的細胞在體外培養時表現出生命周期縮短的表型,而其相關基因則與酵母中編碼蝸牛酶的SGS1基因具有很高的同源性。與來自布盧姆氏綜合徵個體的培養細胞相似,SGS1基因突變的酵母細胞表現出顯著縮短的生命周期。Francoise等研究了170多個通過功能克隆得到的人類基因,發現它們中有42%與酵母基因具有明顯的同源性,這些人類基因的編碼產物大部分與信號轉導途徑、膜運輸或者DNA合成與修復有關,而那些與酵母基因沒有明顯同源性的人類基因主要編碼一些膜受體、血液或免疫系統組分,或人類特殊代謝途徑中某些重要的酶和蛋白質。
模式生物作用
隨著獲得高等真核生物更多的遺傳信息,人們將會發現有更多的酵母基因與高等真核生物基因具有同源性,因此酵母基因組在生物信息學領域的作用會顯得更加重要,這同時也會反過來促進酵母基因組的研究。與酵母相比,高等真核生物具有更豐富的表型,從而彌補了酵母中某些基因突變沒有明顯表型改變的不足。下面將要提到的例子正說明了酵母和人類基因組研究相互促進的關係。人類著色性乾皮病是一種常染色體隱性遺傳的皮膚疾病,極易發展成為皮膚癌。
早在1970年Cleaver等就曾報導,著色性乾皮病和紫外線敏感的酵母突變體都與缺乏核苷酸切除修復途徑(nucleotide excision repair,NER)有關。1985年,第一個NER途徑相關基因被測序並證實是酵母的RAD3基因。1987年,Sung首次報導酵母Rad3p能修復真核細胞中DNA解鏇酶活力的缺陷。1990年,人們克隆了著色性乾皮病相關基因XPD,發現它與酵母NER途徑的RAD3基因有極高的同源性。隨後發現所有人類NER的基因都能在酵母中找到對應的同源基因。重大突破來源於1993年,發現人類xPBp和xPDp都是轉錄機制中RNA聚合酶Ⅱ的TFⅡH複合物的基本組分。於是人們猜測xPBp和xPDp在酵母中的同源基因(RAD3和RAD25)
酵母作為模式生物的作用不僅是在生物信息學方面的作用,酵母也為高等真核生物提供了一個可以檢測的實驗系統。例如,可利用異源基因與酵母基因的功能互補以確證基因的功能。據Bassett的不完全統計,到1996年7月15日,至少已發現了71對人類與酵母的互補基因,這些酵母基因可分為六個類型:
(1)20個基因與生物代謝包括生物大分子的合成、呼吸鏈能量代謝以及藥物代謝等有關;
(2)16個基因與基因表達調控相關,包括轉錄、轉錄後加工、翻譯、翻譯後加工和蛋白質運輸等;
(3)1個基因是編碼膜運輸蛋白的;
(4)7個基因與DNA合成、修復有關;
(5)7個基因與信號轉導有關;
(6)17個基因與細胞周期有關。現在,人們發現有越來越多的人類基因可以補償酵母的突變基因,因而人類與酵母的互補基因的數量已遠遠超過過去的統計。
 釀酒酵母
釀酒酵母在酵母中進行功能互補實驗無疑是一種研究人類基因功能的捷徑。如果一個功能未知的人類基因可以補償酵母中某個具有已知功能的突變基因,則表明兩者具有相似的功能。而對於一些功能已知的人類基因,進行功能互補實驗也有重要意義。例如與半乳糖血症相關的三個人類基因GALK2(半乳糖激酶)、GALT(UDP-半乳糖轉移酶)和GALE(UDP-半乳糖異構酶)能分別補償酵母中相應的GAL1、GAL7、GAL10基因突變。
在進行互補實驗以前,人類和酵母的乳糖代謝途徑都已十分清楚,對有關幾種酶的活性檢測法也十分健全,並已獲得其純品,可以進行一系列生化分析。隨著人類三個半乳糖血症相關基因的克隆分離成功,功能互補實驗成為可能,從而在遺傳學水平進一步確證了人類半乳糖血症相關基因與酵母基因的保守性。人們又將這一成果予以推廣,利用酵母系統進行半乳糖血症的檢測和基因治療,如區別真正的突變型和遺傳多態性,在酵母中模擬多種突變型的組合表型,或篩選基因內或基因間的抑制突變等。這些方法也同樣適用於其它遺傳病的研究。
利用異源基因與酵母基因的功能,還能使酵母成為其它生物新基因的篩查工具。通過使用特定的酵母基因突變株,對人類cDNA表達文庫進行篩選,從而獲得互補的克隆。如Tagendreich等利用酵母的細胞分裂突變型(cdcmutant)分離到多個在人類細胞有絲分裂過程中起作用的同源基因。利用此方法,人們還克隆分離到了農作物、家畜和家禽等的多個新基因。
為了充分發揮酵母作為模式生物的作用,除了發展酵母生物信息學和健全異源基因在酵母中進行功能互補的研究方法外,通過建立酵母最小的基因組也是一個可行的途徑。酵母最小的基因組是指所有明顯豐余的基因減少到允許酵母在實驗條件下的合成培養基中生長的最小數目。人類cDNA克隆與酵母中功能已知基因缺陷型進行遺傳互補可以確定人類新基因的功能,但是這種互補實驗會受到酵母基因組中其它豐余基因的影響。如果構建的酵母最小基因組中所保留的基因可以被人類或者病毒的DNA序列完全替換,那么替換後的表型將完全取決於外源基因,這將成為一種篩選抗癌和抗病毒藥物的分析系統。
盤點鮮為人知的微生物
| 微生物是一切肉眼看不見的或看不清的微小生物的總稱。包括細菌、病毒、真菌以及一些小型的原生動物、顯微藻類等在內的一大類生物群體,它個體微小,卻與人類生活關係密切。 |
