類型
按發生的原因可以把染色體畸變分為自發畸變和誘發畸變。 按畸變的性質可以把染色體畸變分為數目畸變和結構畸變。
數目畸變
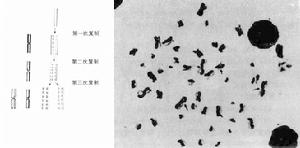 染色體畸變
染色體畸變多數真核生物的體細胞中,都具有兩個染色體組,這樣的生物體和它們的體細胞都稱為二倍體(2n)。二倍體的生殖母細胞經過減數分裂產生的配子中只有一個染色體組,稱為單倍體(n)。某一染色體的數目的增減稱為非整倍性改變;成套的染色體組數目的增減則稱為整倍性改變(見染色體倍性)。非整倍性和整倍性改變統稱為異倍性改變。
一、非整倍性
非整倍性染色體數目畸變可分為:
①單體性二倍體細胞的某同源染色體只有一個而不是兩個的現象,即2n-1。大多數動植物的單體性個體不能存活,存活的單體最初是在小麥中發現的。普通小麥中有成套的21種不同的單體,普通菸草有成套的24種不同的單體,它們是細胞遺傳學研究的有用工具(見基因定位)。在人類中,除特納氏綜合徵(45,X)屬性染色體單體外,常染色體單體性的胚胎常死於子宮內。
 染色體畸變
染色體畸變②缺體性二倍體生物的體細胞缺失了某一對同源染色體的現象,即2n-2。缺體最早在燕麥中發現。缺體性個體一般也不能存活。但普通小麥等少數物種有人工保存的成套缺體性個體。在惡性
腫瘤細胞里也有缺體性細胞系。
③三體性二倍體細胞的某同源染色體為三個的現象,即2n+1。三體的存在最初是在茄科植物曼陀羅中發現的。人類的唐氏綜合徵患者的核型式是47,XX或XY, 21,即21號染色體比正常人多一個。克氏綜合徵患者的核型式是47,XXY,即性染色體X比正常人多一個。三體性個體一般都能存活。
④多體性二倍體細胞的某同源染色體的數目在三個以上的現象。例如人類染色體病中見到的48,XXXX四體或49,XXXXX五體等都是。
超二倍性和亞二倍性是指二倍體生物的體細胞多了若干個或少了若干個染色體的現象,也屬於非整倍性畸變,常見於離體培養的腫瘤細胞。
 染色體畸變
染色體畸變產生非整倍性個體的主要原因是在生殖母細胞經減數分裂形成配子的過程中或在受精卵的卵裂過程中染色
體的複製和分配異常所致,而且主要是由減數分裂後期Ⅰ某一對同源染色體不分離或後期Ⅱ某一染色體的兩個染色單體不分離引起的。如果不分離發生在配子形成過程中,那么就形成n 1和n-1兩類異常的配子。當這些配子與正常配子(n)結合,就發育為單體性(2n-1)或三體性(2n 1)個體。如果不離開發生在受精卵經卵裂形成早期胚胎的過程中,那么單體性和三體性的體細胞就可以在同一個體中同時存在,從而形成嵌合體。
二、整倍性 見染色體倍性。
結構畸變
染色單體畸變 染色單體或染色單體間結構的變化有兩種形式:①簡單的缺失,即單體斷裂下來的片斷丟失;②結構重排,即發生在同一染色體臂內或臂間的單體內互換和發生在不同染色體的單體間的互換。互換可以相等或不相等。 單體間互換按重接方式又可分為兩種類型。如果斷裂端以著絲粒為中心的近心部與近心部相接,遠心部與遠心部相接,稱為U型互換。如果斷裂端的近心部與遠心部相接,則稱為X型互換。U型互換是不對稱互換,X型互換是對稱互換。
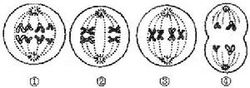 染色體畸變
染色體畸變同一染色體發生的臂間和臂內互換可以有6種方式,而不同染色體單體間的互換則根據配對的同源染色體、互換的類型、互換是否完全以及染色體的極性又可分為12種情況。
除了簡單的單體內互換和單體間互換外,有些畸變是由於在染色體和染色單體水平上發生多次互換而產生的,例如三相互換。這種畸變的產生是由於在一個等位點染色單體畸變和一個簡單的染色單體斷裂之間發生了一次互換,或者在兩個以上染色單體之間發生數次複雜的單體間互換造成的。
染色體的結構變化主要有以下幾種:
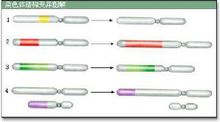 染色體畸變
染色體畸變①缺失染色體臂發生斷裂並丟失一部分遺傳物質的結果。一個染色體臂發生了斷裂,而這種斷裂端未能與別的斷裂端重接,那么就形成一個帶有著絲粒的片段和一個沒有著絲粒的片段。後者在細胞分裂過程中
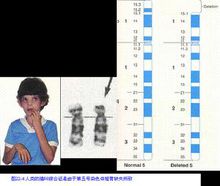
不能定向而被丟失。帶有著絲粒的片段便成為一個發生了末端缺失的染色體。如果一個染色體發生兩次斷裂而丟失了中間不帶有著絲粒的片段,留下的兩個片段重接以後便成為發生了中間缺失的染色體。如果同一染色體的兩臂同時發生了斷裂,而餘下的兩臂的斷面間又發生重接,便形成環形染色體。根據所丟失的染色體片段的大小,缺失所帶來的危害性也各不相同。較大的缺失往往帶來致死效應,而微小的缺失則並不致死。如果缺失部分包括某些顯性等位基因,那么同源染色體上與這一缺失相對應位置上的隱性等位基因就得以表現,這一現象稱為假顯性。在玉米中,如果染色體上帶有顏色決定基因的區段缺失則常能產生特定的表型效應,例如白苗與褐色中脈等。在人類中,染色體的部分缺失常導致染色體病,如貓叫綜合徵就是由於5號染色體的短臂部分缺失所致。
 染色體畸變
染色體畸變②重複一個染色體上某一部分出現兩份或兩份以上的現象。首尾相接的重複稱為銜接重複或串接重複;首尾反方向連線的重複稱為顛倒銜接重複或倒重複。重複部分可以出現在同一染色體上的鄰近位置,也可以
出現在同一染色體的其他位置或者出現在其他染色體上。重複雜合體具有特徵性的減數分裂圖象,它的染色體在進行聯會時重複片段在同源染色體上找不到相應的結構,因而形成稱為重複環的環狀突起。類似的圖象可以在果蠅的重複雜合體的唾腺染色體中看到。在缺失雜合體細胞中也同樣可以看到圖象相似的缺失環。重複的遺傳效應比缺失來得緩和,但重複太大也會影響個體的生活力,甚至引起個體的死亡。染色體上某些區域的重複可以產生特定的表型效應,例如果蠅的顯性基因棒眼(Bar eye,B)就是重複的結果。主要的表型效應是複眼中的單眼數減少,使複眼呈棒狀而不是通常的卵圓形。在這種果蠅的唾腺染色體上可以看到X染色體上明顯的橫紋重複(見位置效應)。可是對於一般的染色體來說,不通過顯帶法是很難檢出重複的。
③倒位 一個染色體上同時出現兩處斷裂,中間的片斷扭轉180°,重新連線起來而使具有同源染色體的細胞這一片段的基因的排列順序顛倒的現象。顛倒片段包括著絲粒的倒位稱為臂間倒位;不包括著絲粒的倒位稱為臂內倒位。兩個斷裂點與著絲粒之間的距離不等的臂間倒位是容易識別的,等距離的倒位則除非套用顯帶技術一般不易察覺。倒位雜合體也具有特徵性的減數分裂圖象,它的倒位染色體和正常同源染色體聯會時出現倒位環。。臂內倒位雜合體如果在倒位環內發生一次交換便形成一個有兩個著絲粒的染色單體和一個沒有著絲粒的片斷,這樣在減數分裂後期就會出現染色體橋和沒有著絲粒的片段,後者往往無法進入子細胞核中;而有兩個著絲粒的橋被拉斷後,雖然兩個染色體可分別進入子細胞,但由於斷裂位置不一,往往帶來缺失而導致配子的死亡。
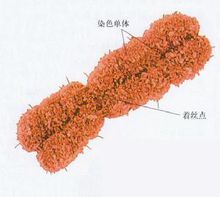 染色體畸變
染色體畸變④易位一個染色體臂的一段移接到另一非同源染色體的臂上的結構畸變。兩個非同源染色體間相互交換染色體片段稱為相互易位。相互易位的染色體片段可以是等長的,也可以是不等長的。一般基因改變它在染色體上的位置時並不改變它的功能,可是在果蠅等生物中發現如果位置在常染色體的基因通過易位而處於異染色質近旁時,它的功能便會受到影響而呈現花斑位置效應現象。倒位也可能帶來同一效應。易位純合體沒有明顯的細胞學特徵,它在減數分裂時的配對也不會出現異常,因而易位染色體可以從一個細胞世代傳到另一個細胞世代。可是易位雜合體則不同,由於正常染色體和易位染色體的同源部分的配對,在減數分裂中期可以看到特有的十字形圖象。 隨著分裂過程的進行,十字形逐漸開放,鄰接的兩個著絲粒趨向同一極或趨向兩極,形成環形或8字形圖象。前一種染色體離開方式稱為鄰近離開,後一種方式稱為互動離開。相互易位雜合體的花粉母細胞中大
約有50%的圖象呈環形,屬鄰近離開,50%呈8字形,屬互動離開。這說明四個著絲粒趨向兩極的取向是隨機的,行動是獨立的。鄰近離開的結果使配子內含有重複或缺失的染色體,形成致死的不平衡配子。互動離開導致非致死的平衡配子,其中半數配子的染色體是正常的,半數配子具有平衡的易位染色體,這就是說雖然發生了易位,可是易位並不帶來基因的增加或缺少。互動離開使兩個易位染色體進入一個配子細胞,兩個非易位染色體進入另一配子細胞中。所以這種分離方式使非同源染色體上的基因間的自由組合受到限制,使原來在不同染色體上的基因出現連鎖現象,這種現象稱為假連鎖(見基因定位)。
兩個或兩個以上染色體之間發生相互易位時,如果這些染色體的近心端相接,就形成雙著絲粒體或多著絲粒體。雙著絲粒體同時具有兩個有功能的著絲粒,它們在細胞分裂後期各自趨向兩極,從而出現後期橋。如果這種橋被拉斷,往往導致細胞死亡。然而根據玉米胚乳細胞的研究,認為染色體橋雖可被拉斷,但兩端的兩個著絲粒仍可順利地進入細胞的兩極並參與子細胞的形成,而且它們的斷裂端仍然開放著並可重接。當再次相接後就會在下一次分裂的後期形成另一個橋,這種“斷裂-融合-橋”周期可以持續許多個細胞世代。
整臂易位是整個臂(或幾乎是整個臂)之間的易位,這種易位的結果可以產生結構不同的兩個新的染色體。在整臂易位中又有一種特殊的情況,即兩個同源的(或非同源的)近端著絲粒染色體的著絲粒相互融合,成為一個中央(或亞中央)著絲粒染色體,結果導致染色體數目減少,但臂數不變。這種整臂易位稱為羅伯遜氏易位,為W.R.B.羅伯遜於1916年所發現。例如小鼠屬中,最普遍的核型是40個近端著絲粒染色體,可是在某些野鼠的細胞中則出現若干雙臂染色體。這些雙臂染色體就是通過著絲粒融合形成的。一般認為在哺乳動物的核型進化中,著絲粒融合是最普通的一種形式。
⑤ 環狀染色體(ring):如染色體的長、短臂同時各發生一次斷裂後,含有著絲粒節段的長、短臂斷端相接,即形成環狀染色體。這一異常在遺傳上是不穩定的,因為其染色體環隨染色體複製而必須打開一次。
⑥ 等臂染色體(isochromosome):當染色體的著絲粒非以縱向分裂,而發生了橫向分裂時,使一個子代細胞接受兩條長臂,另一個接受兩條短管所形成之。等臂染色體是最常見的染色體的結構異常。
⑦ 染色體複製(duplication):系染色體的一部分被複製,新複製的一段染色體可位於同條染色體內,或附著到另一條染色體上,或成為獨立的節段。實際上,染色體複製的發生率高於染色體缺失,但由於並無遺傳物質的丟失,表型異常不常見,而可能被忽視之。
除了上述幾種結構的畸變以外,在光學顯微鏡下還可以看到染色體的另外一些形態上的變化。例如染色體粘連,粘連在一起的染色體數目可以是兩個或兩個以上。在細胞有絲分裂和減數分裂過程中許多理化因素可誘發染色體粘連,而某些突變基因也能促使染色體發生粘連。此外,環境因子的劇烈作用還可以有種種效應:染色體粉碎化(高頻度的斷裂)、單體化(中期染色體以單體形式存在)、不同步化(同一細胞內染色體的形成速度不一致)、解鏇化(中期染色體被解螺鏇而鬆開)等。
引發疾病
先天愚型(21三體綜合徵)
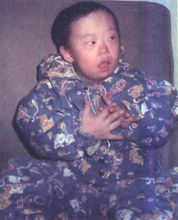 染色體畸變
染色體畸變患兒具有特殊的顱面部畸形,頭顱小而圓,枕骨扁平,眼小而眼距過寬,兩眼外側高而內側低,臉圓,鼻扁平,口半張,舌常伸出口外,舌有龜裂,低耳位,常為通貫手,發育遲緩,智力低下,平均壽命很短,大約到10歲時已有1/3患者死亡。病因是因為多了一條小的第21號染色體。患者大多都同時患有先天性心臟病。
Patau綜合徵(13三體綜合徵)
具有額外的13號染色體,記作47,XX或XY, 13,發病率約為1/5000。患者中女性明顯多於男性,患兒的畸形和其他臨床特徵比21三體要嚴重,小頭、兔唇和(或)齶裂、先天性心臟病、嚴重智力遲鈍。90﹪在出生後6個月內死亡。
Edwards綜合徵
由於有額外的18染色體(E組)所致核型為47,XX或XY, 18。另有8﹪—10﹪的病例為48,XXX或XXY, 18,雙重非整倍性。這類患兒不僅有額外的18號染色體,還有一條額外的X染色體。它會導致嚴重的畸形,在出生後不久死亡,平均年齡僅為71天。因為其畸形幾乎遍及所有的器官系統,此外95﹪的患兒有先天性心臟病,是嬰兒致死的重要原因。
貓叫綜合徵
5號染色體的缺失造成的,患者幼兒時期,哭聲好像貓叫。故稱貓叫綜合徵。
兩種假說
斷裂-重接假說
由L.J.斯塔德勒於1931年提出。另一種是互換假說,由S.H.雷維爾於1959年提出。 斷裂-重接假說認為導致染色體結構改變的原發損傷是斷裂。這種斷裂可以自發地產生,也可以是誘變因子作用的結果。斷裂的後果不外是以下三種:①絕大多數斷裂(90~99%)通過修復過程在原處重新連線(癒合)以致在細胞學上無法予以辨認。②不同斷裂處的重新連線稱為重接,重接使染色體發生結構變化,所以大多能被發現。③斷裂端依然游離著,成為染色體結構的一種穩定狀態,例如末端部分缺失。
互換假說
認為導致染色體結構畸變的根本原因是染色體上具有不穩定部位,所有結構畸變都是兩個靠得很近的不穩定部位之間互換的結果。互換的發生可以分為兩個階段:第一階段是不穩定部位斷裂後繼發的較穩定狀態,稱為互換起始。第二階段是機械的互換和連線過程。如果兩個原發的損傷不能相互作用,這些損傷就可以被修復。
