主要種類
 同源基因
同源基因1.直向同源基因
直向同源基因(orthologous gene)又譯為“垂直同源基因”、“正同源基因” 或“定向進化同源基因”、“直系同源基因”,是指從同一祖先垂直進化而來的基因。或者說,一個祖先物種分化產生兩種新物種,那么這兩種新物種共同具有的由這個祖先物種繼承下來的基因就稱為直向同源基因。直向同源基因通常是編碼生命必需的酶、輔酶或關鍵性的調控蛋白的基因,具有功能保守,進化緩慢,變化速度可覆蓋整個進化歷史,且序列變化速度與進化距離相當等特徵。大多數直向同源基因功能相同或相近,調控途徑也相似, 因此在基因組序列的注釋中,是最可靠的選擇,例如人α一珠蛋白基因與小鼠α一珠蛋白基因。
2.橫向同源基因
橫向同源基因(paralogous gene)又譯為“旁系同源基因”、“並系同源基因”或“平行進化同源基因”,是指由於基因重複而產生的同源基因例如人γ一珠蛋白基因和β一珠蛋白基因。基因重複後,進化選擇壓力變小、其中一條基因丟失或發生沉默都是促使橫向同源基因分化產生新特性或新功能的原因。然而,雖然某些橫向同源基因轉錄區序列相似度不高,但它們的操縱子卻仍然具有較高的保守度。值得注意的是,橫向同源基因並不局限於同一物種內,不同物種中由於始祖基因的複製而分化的基因也稱橫向同源基因,如鼠α一珠蛋白和雞β一珠蛋白基因。
3.異源同源基因
異源同源基因(xenologous gene)是由於基因在不同物種間的橫向轉移(horizontal transfer)而產生的。異源同源基因在原核生物中研究比較多。最近研究表明,異源同源基因的原位取代xenolo—gous gene displacement in situ)是細菌進化的強大推動力。另外,在比較真核基因組和原核生物基因組時發現,小部分脊椎動物基因在細菌中有同源序列,而在其他真核生物中卻沒有發現同源序列。一種解釋認為,這些基因從細菌直接水平地轉移到脊椎動物的祖先,也是異源同源基因;另外一種解釋則認為,是由於其他的真核生物丟失了這些基因。
相互聯繫
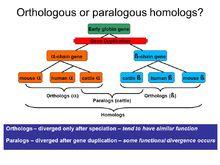
基因的直向同源、橫向同源或異源同源關係見圖1。如圖1所示,祖先物種通過兩次物種分化形成ABC三個物種;伴隨物種分化而進行的兩次基因重複共形成A1、B1、B2、C1、C2、C3等6個基因。顯然,C2與C3互為橫向同源;B1與C1互為直向同源;AB1與其他6個基因互為異源同源。然而,B1和B2、B2和C1又是什麼關係呢?在這個問題上曾引起爭議。我們從圖中可以看出,B1和B2、B2和C1的分離是由於第一次基因重複而產生的,套用定義,可得出B1和B2、B2和C1互為橫向同源。同理,B1和C2/C3、C1和C2/C3也互為橫向同源。類似的,根據直向同源基因的定義,B2和C2/C3、A1和所有B基因及C基因互為直向同源。
以上只是根據基本定義進行的大致的歸分。然而,在研究親緣關係較遠的物種時,物種分化和基因重複的嵌套發生使研究具有很大的複雜性和困難度。這時,籠統的劃分直系同源和旁系同源關係並不足以解決問題,我們需要一個更為精確的分類。在圖1中,為了區別於B1和C1的簡單的直向同源關係,把B2和C2/C3的同源稱為“共生直向同源”co—ortholog)或“橫向同源基因家族譜系特異性擴充”(1ineage—specific expansion ofparalogous family)。為了區分C2和C3、B1和C2/C3這兩種同源關係,可以根據物種分化和基因重複的發生的先後次序,把橫向同源基因又分為內橫向同源基因(inparalog)和外橫向同源基因(outparalog)。在物種分化之後產生橫向同源基因的稱“內橫向同源基因”;在物種分化之前產生橫向同源基因的稱“外橫向同源基因”。 “內橫向同源基因”和“外橫向同源基因” 只是一種相對的劃分,取決於所研究的系統和選作標準的某特定分化事件。圖1中,若以第二次物種分化為準線,則C2和C3的分離發生在物種分化之後,因此它們互為內橫向同源基因;B1和C2/C3的分離發生在物種分化之前, 因此B1和C2/C3互為外橫向同源基因。
