
概述
 珊瑚礁的形成過程
珊瑚礁的形成過程簡介
珊瑚礁是石珊瑚目的動物形成的一種結構。這個結構可以大到影響其周圍環境的物理和生態條件。在深海和淺海中均有珊瑚礁存在。它們是成千上萬的由碳酸鈣組成的珊瑚蟲的骨骼在數百年至數千年的生長過程中形成的。珊瑚礁為許多動植物提供了生活環境,其中包括蠕蟲、軟體動物、海綿、棘皮動物和甲殼動物。此外珊瑚礁還是大洋帶的魚類的幼魚生長地。
 美麗的珊瑚礁
美麗的珊瑚礁形成
其形成是造礁珊瑚及其他造礁生物對生成礁的鈣物質長期積累沉積的結果,由造礁珊瑚的石灰質遺骸和石灰質藻類堆積而成的一種礁石。世界上珊瑚礁多見於南北緯30°之間的海域中,尤以太平洋中、西部為多。
形態
按形態劃分有:裾礁(岸礁)、堡礁、環礁、桌礁及一些過渡類型。據估計全世界珊瑚礁連同珊瑚島面積共有1000萬平方公里。珊瑚礁生長速度一般為每年2.5厘米左右。有些珊瑚礁厚度很大,系因珊瑚礁生長發育過程中礁基不斷下沉或海面不斷上升所致。達爾文根據礁體與岸線的關係,劃分出岸礁、堡礁和環礁,根據形態分出台礁和點礁等類型。
組成
珊瑚礁是由石珊瑚目的珊瑚蟲的骨骼組成的。這些骨骼的主要成分是碳酸鈣(CaCO3)。珊瑚的端部不斷成長在死去的珊瑚蟲的骨骼上。海浪、遊動的魚和其他力以及生物折斷這些珊瑚,它們落入珊瑚礁的隙縫中。許多其他生活在珊瑚礁的生物的骨骼也是由碳酸鈣組成的,它們也為珊瑚礁的形成做出貢獻,但是珊瑚蟲是最重要的。尤其海浪比較強的地方珊瑚蟲所造成的結構是最主要的。它們一層一層地加厚珊瑚礁,為珊瑚礁提供了其結構強度。其他珊瑚在珊瑚礁的表面形成樹枝似的結構,加大了珊瑚礁的面積。甲殼動物進一步加強了珊瑚礁的強度,防止它被海浪摧毀。這些甲殼往往在珊瑚礁的邊緣形成一個保護層。這個結構尤其在太平洋非常顯著。
生長環境
大多數石珊瑚必須在大洋的透光層(水深不到50米)中生長,這裡珊瑚內部共生的單細胞的蟲黃藻能夠進行光合作用。這些蟲黃藻通過光合作用來為珊瑚蟲提供營養。因此珊瑚礁在清晰的水中的生長速度最高。事實上,沒有共生的蟲黃藻珊瑚礁的生長速度非常慢,不可能達到其客觀的結構。
一般珊瑚礁長到水面就不繼續生長了。原因是大多數珊瑚蟲不能在水面上生長,少數可以在水面上生長的珊瑚蟲也不能長時間脫離水。珊瑚礁最主要的生長區位於其邊緣,這裡的珊瑚礁一般緩慢下降,然後在礁石的邊緣突然陡降。珊瑚蟲的生長也幫助產生水流來運入無機營養,運走新陳代謝產物。珊瑚礁的邊緣同時也是受風浪衝擊最強的地方。這裡生長與破壞形成了一種平衡。被風浪折斷的珊瑚礁落入深處,逐漸加闊珊瑚礁的地基。
珊瑚礁錯綜複雜的結構為許多魚和無脊椎動物提供了多樣的生活環境。
資源
珊瑚從古生代初期開始繁衍,一直延續至今,可作為劃分地層、判斷古氣候、古地理的重要標誌。珊瑚礁與地殼運動有關。正常情況下,珊瑚礁形成於低潮線以下50米淺的海域,高出海面者是地殼上升或海平面下降的反映;反之,則標誌該處地殼下沉。珊瑚礁蘊藏著豐富的油氣資源。珊瑚礁及其潟湖沉積層中,還有煤炭、鋁土礦、錳礦、磷礦,礁體粗碎屑岩中發現有銅、鉛、鋅等多金屬層控礦床。珊瑚灰岩可作燒石灰、水泥的原料,千姿百態的珊瑚可作裝飾工藝品,不少礁區已開闢為旅遊場所。
形成過程
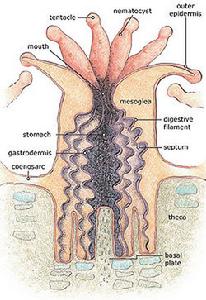 珊瑚蟲
珊瑚蟲珊瑚礁的主體是由珊瑚蟲組成的。珊瑚蟲是海洋中的一種腔腸動物,它以捕食海洋里細小的浮游生物為食,在生
長過程中能吸收海水中的鈣和二氧化碳,然後分泌出石灰石,變為自己生存的外殼。每一個單體的珊瑚蟲只有米粒那樣大小,它們一群一群地聚居在一起,一代代地新陳代謝,生長繁衍,同時不斷分泌出石灰石,並粘合在一起。這些石灰石經過以後的壓實、石化,形成島嶼和礁石,也就是所謂的珊瑚礁。由於珊瑚蟲具有附著性,許多珊瑚礁的底部常常會附著大量的珊瑚蟲。珊瑚是非生物,屬刺胞動物門,當中也包括水母、水螅、軟珊瑚、海葵等動物。珊瑚由很多珊瑚蟲造成。每一珊瑚蟲都有一個中空而底部密封的柱型身體,它的腸腔與四周的珊瑚蟲連線,而位於身體中央的口部,四周長滿觸手我們通常把珊瑚分為石珊瑚,八放珊瑚及水螅珊瑚,它們有不同的形態特徵。除了生物學分類外,我們亦可按生態功能,把珊瑚分為兩大組。那些有共生藻(即蟲黃藻)的珊瑚稱為可造礁珊瑚,而那些沒有共生藻的則稱為不可造礁珊瑚。
研究簡史
 珊瑚礁
珊瑚礁早在225~230年,中國三國(吳)時期,康泰在《扶南傳》中記載了南海的珊瑚礁。19世紀初,德國自然學者A.von沙米索等在印度洋航行中,發現一些低矮群島是由坐落於海底山頂上的造礁珊瑚構成,並指出礁體形狀與盛行風和水流等有關。1831~1836年,進化論創始人C.R.達爾文乘“貝格爾”艦環球考察中,對珊瑚礁進行過觀察,劃分出岸礁、堡礁和環礁,並於1842年在《珊瑚礁的構造和分布》一文中,提出了珊瑚礁成因的“沉降說”,極大地推進了珊瑚礁的研究。有關其成因的討論以後持續了近100年。1912年法國L.儒班發表了第一幅全球珊瑚礁分布示意圖。
1935~1937年,中國地質學家馬廷英利用珊瑚的生長紋與骨骼密度差關係,測得中國東沙群島造礁珊瑚成長率每年為4~11毫米。50年代以來,美國H.S.萊德、K.O.埃默里和J.W.韋爾斯等人發現珊瑚礁主要分布在受赤道暖流明顯影響的大洋西部。1957年,韋爾斯闡明了珊瑚礁發育主要受水溫、鹽度、水深和光照等因素控制。1968~1970年間,澳大利亞學者W.G.H.馬克斯韋爾對昆士蘭大堡礁作了詳細研究,闡明了水文條件變化與礁體形態和發展的關係;他所提出的珊瑚礁分類,是對達爾文的分類的重要補充。20世紀40年代以來,在古代礁中發現了不少大型油氣田。由此更促進了珊瑚礁研究工作的開展,並使現代礁與古代礁的研究結合起來。
海水溫度上升將殺死珊瑚礁
2010年1月14日,生態保護組織世界自然基金會在一份報告中表示,如果再不採取有效措施,東南亞珊瑚礁的生物多樣性將在本世紀末消失,摧毀沿岸經濟。世界海洋大會在蘇拉威西的馬那多召開。這份報告正好在開會時出爐。會上指出,袖手旁觀將放任海水溫度穩步上升,並殺死珊瑚礁。
形成環境
 珊瑚礁
珊瑚礁造礁珊瑚對水溫、鹽度、水深和光照等條件都有比較嚴格的要求:
①水溫。珊瑚生長的水溫約為20~30°C。J.D.米利曼認為23~27°C是造礁珊瑚生長發育的最佳水溫,韋爾斯認為最佳水溫上限可達29°C。熱帶海區,這一最佳水溫出現在冬季和春季,因而許多學者認為冬季珊瑚生長最快。海南島和西沙群島水溫平均為25~27°C,屬珊瑚生長最佳水溫範圍,但海南島的季節變化大,水溫不穩定,對珊瑚生長有抑制作用。海南島和台灣的珊瑚礁被稱為“高緯度珊瑚礁”。
②鹽度。造礁珊瑚生長在鹽度為27~40的海水中,最佳鹽度範圍是34~36。南海鹽度為34,屬最佳鹽度範圍,海南島沿岸有淡水注入,鹽度略低,為32左右。
③水深。一般認為造礁珊瑚生長的水深範圍是0~50米,最佳水深為20米以淺。許多學者認為這實際上是與造礁珊瑚共生的蟲黃藻進行光合作用所需的深度。
④光照。也是蟲黃藻光合作用的需要,一般熱帶光照強,時間長,平均光照率在50%以上,有利於珊瑚礁的發育。
⑤風和風浪。一般迎風浪一側礁發育較好。新月形和馬蹄形礁體的凸面是迎風迎浪的。如果風浪有季節性變化,礁的形狀會出現雙馬蹄形。所以根據古代礁的形態可判斷古風向。過強的風浪使珊瑚蟲難以在基底上固著,不發育礁。
⑥河流。河流入海處,海水鹽度低,泥沙含量大,混濁度高,海水透明度低,會使珊瑚窒息而死,所以有大量泥沙入海的河口處一般不發育岸礁,如海南島的岸礁在河口區缺失。
 珊瑚礁的形成過程
珊瑚礁的形成過程⑦海平面變動。當海面穩定時,珊瑚礁平鋪發展,但厚度不大;當海面上升或海底下沉時,形成的礁層厚度較大,礁體可發育成塔形、柱形,也有的礁體可深溺於海面以下成為溺礁。當海面下降或地殼上升時,形成的礁層厚度也不大,也有的礁體可高出海面成為隆起礁。這種影響因素對古代礁意義較大。
⑧海底地形和底質。無論在大洋或淺海區,珊瑚礁總是生長於海底的正地形上,如大洋中的平頂海山、海底火山、大陸架的邊緣堤以及構造隆起上。由於在不同的海底地形上水動力作用不盡一致,因此地形特徵有時對礁體發育有很大影響。如極淺的平緩海底往往形成離岸礁;而岸坡較陡,則礁體緊貼岸線發育。珊瑚在海底營固著生活,在堅硬的岩石基底上發育較好,部分屬種也可在水下砂坎上發育,說明對底質有一定的選擇。
⑨藻類與珊瑚礁的關係。蟲黃藻與造礁珊瑚共生,它吸收造礁珊瑚排出的CO2,為珊瑚蟲提供鈣質,形成骨骼中甲殼質(幾丁質)的有機成分,它們構成一個相互依存的生態系統。紅藻中的珊瑚藻是完全鈣化藻,可形成層狀骨架,參與造礁。藻屑是珊瑚礁中常見的組分,一般占20~50%。藻類還可粘結礁骨架和生物屑,並有富鎂作用,形成高鎂方解石。但鑽孔藻(Bringalgal)在珊瑚礁中起破壞作用。
生物多樣化
儘管珊瑚礁位於營養少的熱帶海洋中,它們的生物多樣性非常高。僅珊瑚蟲與蟲黃藻之間的營養循環就可以體現出這個高多樣化的原因:通過回收只有很少的營養會流失到整個生態環境外。
藍藻為珊瑚礁提供可溶的營養物質。珊瑚蟲直接從海水吸收營養,包括無機氮和磷。此外它們吃被水流帶到它們附近的浮遊動物。因此珊瑚礁的初級生產量非常高。珊瑚礁的生產者還包括與珊瑚蟲共生的蟲黃藻和不同的海帶。對於比較小的藻類的作用在科學界有爭議。
許多熱帶或者珊瑚魚住在珊瑚礁內,比如色彩鮮艷的鸚嘴魚、琪蝶魚、雀鯛、蝴蝶魚。其他在珊瑚礁中生存的魚類包括石斑魚、笛鯛、石鱸、隆頭魚等等。至今在珊瑚礁一共發現了4000多種魚。
許多其他生物也以珊瑚礁為家,其中包括海綿、腔腸動物、蠕蟲、甲殼動物、軟體動物、棘皮動物、尾索動物、海龜、海蛇等。除人外哺乳動物很少在珊瑚礁活動。偶爾到珊瑚礁的海豚是最主要的哺乳動物。所有這些動物有的直接吃珊瑚,其他的吃藻類、水草等。它們構成一個複雜的食物網[8]。
一些無脊椎動物生活在珊瑚礁的岩石基礎內部。有些能夠鑽入石灰岩,有些住在岩石內部本來就存在的縫隙中。能夠在石灰岩中打洞的動物包括海面、雙殼軟體動物和星蟲動物。居住在原有洞穴中的動物包括甲殼動物和沙蠶。
許多國家政府因此保護它們的珊瑚礁。比如澳大利亞設立了大堡礁海洋公園來保護大堡礁,其措施包括立法和行政計畫,包括生物多樣性行動計畫。
分布
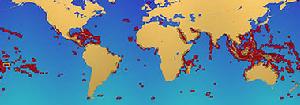 珊瑚礁的位置
珊瑚礁的位置珊瑚礁的位置全世界的珊瑚礁總面積估計為28.43萬平方公里,其中印度洋-太平洋地區(包括紅海、印度洋、東南亞和太平洋)占91.9%的面積。僅東南亞就占32.3%的面積,太平洋(包括澳大利亞)占40.8%。大西洋和加勒比海僅占全世界的7.6%。
美國西海岸和非洲西海岸基本上沒有珊瑚礁,或者很少,其原因主要是上升的牆冷海流降低當地的水溫。從巴基斯坦到孟加拉國的南亞海岸的珊瑚礁也很少。南美洲東南海岸和孟加拉國缺少珊瑚礁的原因是因為亞馬孫河和恆河在這裡有大量淡水入海。
世界上著名的珊瑚礁有:
澳大利亞的大堡礁是世界上最大的珊瑚礁
中美洲宏都拉斯的羅阿坦堡礁是世界上第二大的珊瑚礁
埃及紅海海岸的珊瑚礁
種類
按照其地理分布的不同珊瑚礁可以分兩類:深水珊瑚礁和熱帶珊瑚礁。
深水珊瑚礁
許多石珊瑚也可以在水溫20°C以下成長和成礁。不像熱帶的珊瑚,它們不使用陽光作為首要能源,而是使用周圍水裡的營養。與熱帶的珊瑚礁相比這些珊瑚礁的生長非常緩慢。比如在歐洲的沿海從伊比利亞半島直到北角有一圈這樣的珊瑚礁圍繞。形成這些珊瑚礁的主要物種是Lophelia pertusa和Madrepora oculata。
典型的珊瑚礁位於水深200至1000米之間,至今為止被發現的最深的深水珊瑚礁達3000米。40米以上還沒有發現有屬於這些屬的珊瑚生存。
深水珊瑚礁尤其受到現代化的捕魚技術的威脅。深水拖網可以在幾分鐘內破壞整個在數千年里緩慢成長起來的珊瑚礁。
熱帶珊瑚礁
水溫常年高於20°C的地區熱帶珊瑚礁的珊瑚只能在水溫高於20°C的地區生存。這些石珊瑚與蟲黃藻共生,因此它們需要充分的陽光,它們因此生活在水深50米以上的水中。因此熱帶珊瑚礁一般位於北緯30°至南緯30°之間。熱帶珊瑚礁的總面積約為60萬平方公里,它們每年約堆積6.4億噸的碳酸鹽。
熱帶珊瑚礁分兩類:
沿岸沉積形成的珊瑚礁位於大陸架的淺水中。由於淡水帶來的營養物質這些珊瑚礁營養比較豐富。因此這些珊瑚礁中往往以軟珊瑚和藻類植物為主。
火山島形成的珊瑚礁遠離大陸,它們位於由火山活動形成的島嶼的周圍。比如夏威夷和塔希提周圍的珊瑚礁。這些珊瑚礁的營養比較少,因此以石珊瑚為主。
形態
 珊瑚礁
珊瑚礁按照其形式珊瑚礁可以分多個形態。
環礁
環礁一般是由火山島周圍的裾礁演化而成的。通過風化島嶼逐漸被消磨,最後沉到水面以下,最後只剩下一個環繞著一個暗礁的環礁。海底下沉和海面上升也會形成環礁。馬爾地夫由26個這樣的環礁組成。
裙礁
裙礁沿著大陸或者島嶼的邊緣形成。大多數沿岸珊瑚礁是裙礁。裙礁的形成需要一定的條件,在大量淡水入海的地方(比如亞馬孫河的入海口)裙礁無法形成。塞席爾是一個遠洋裙礁的例子。
平頂礁
假如海底有平坦的、珊瑚礁生長的條件,那么就能夠形成平頂礁。平頂礁有時有些地方可以伸出水面,在那裡形成沙灘和小島,而在這些沙灘和小島附近又可以形成裙礁。在平頂礁的中部可能形成礁湖。環礁的內部也可能有平頂礁形成。
 珊瑚礁的形態
珊瑚礁的形態堡礁
堡礁位於大陸架的邊緣,它在大洋與大陸架的淺水之間形成了一個屏障。堡礁可以是因為大陸下沉由裙礁演化而成。最著名的堡礁是澳大利亞的大堡礁。
達爾文根據礁體與岸線的關係,劃分出岸礁、堡礁和環礁,根據形態還有台礁和點礁等類型。
①岸礁。沿大陸或島嶼岸邊生長發育,亦稱裙礁或邊緣礁。現代最長的岸礁沿紅海沿岸發育,綿延約2700多公里,分布水深約36米。中國台灣恆春半島和海南島沿岸也有岸礁發育。
②堡礁。又稱堤礁,是離岸有一定距離的堤狀礁體,它與陸地隔以瀉湖。現代規模最大的堡礁是澳大利亞昆士蘭大堡礁,全長約2000公里,分布水深約30米。
③環礁。礁體呈環帶狀圍繞瀉湖,有的與外海有水道相通。環礁直徑在幾百米至幾十公里,形態多樣。已知的環礁有330個之多,主要分布在西太平洋的信風帶和印度洋熱帶海域。環礁多坐落在大洋火山錐上,孤立於汪洋大海之中,展布受洋底火山作用的控制,某些也可在大陸架上見到。環礁礁坪上常有灰砂(礫)島或礁岩島,統稱為珊瑚島。馬紹爾群島上的夸賈林環礁和馬爾地夫群島的蘇瓦迪瓦環礁,面積都在1800平方公里以上,是世界上最大的兩個環礁。南海發育的環礁頗具特色,有瀉湖全被封閉的玉琢礁;有瀉湖與外海有3個通道的華光礁;還有多通道開放式的永樂環礁,半月形全開放式的宣德環礁。永樂環礁和宣德環礁均發育有灰砂島和礁岩島,其中最大的灰砂島是永興島,面積1.85平方公里。
④台礁。呈台地狀高出附近海底,但無瀉湖和邊緣隆起的大型珊瑚礁。也稱桌礁。礁坪上也可發育灰砂島,如西沙群島的中建島。
⑤點礁。即斑礁,是堡礁和環礁瀉湖中的礁體,大小不等,形態多樣。
根據形態還有圓丘礁、塔礁、馬蹄礁、層狀礁等。馬克斯韋爾將昆士蘭大堡礁的礁體分成月牙礁、耙狀礁和封閉網狀礁等19個類型。礁體往往成群展布。根據礁群特徵又可分為:線型礁群、寬頻狀礁群、弧形或馬蹄形礁群和環形礁群等。根據礁與海平面的關係,又可分為上升礁和溺礁。上升礁又稱隆起礁。它出露于海面以上,礁上沒有活珊瑚生長,是地殼上升或海面下降的產物。溺礁的形成是因為地殼下沉或海面上升迅速,造礁珊瑚的生長追隨不上,致使礁體沉溺於造礁珊瑚生長的極限深度以下。這裡活的珊瑚無法生存,珊瑚礁遭受溶蝕。因而,溺礁常常被作為地殼下沉或海面上升的可靠標誌。上升礁如台灣和海南島南岸所見,溺礁以中沙群島大環礁為代表。
分類
石珊瑚目 Scleractinia Bourne, 1900
蜂巢珊瑚科 Faviidae Gregory, 1900
扁腦珊瑚屬 Platygyra Ehrenberg
精巧扁腦珊瑚 Platygyra daedalea (Ellis & Solander)
扁腦珊瑚 Platygyra sp
蜂巢珊瑚屬Favia Oken
標準蜂巢珊瑚 Favia speciosa (Dana)
角蜂巢珊瑚屬Favites Link
多彎角蜂巢珊瑚 Favites flexuosa (Dana)
秘密角蜂巢珊瑚 Favites abdita (Ellis & Solander)
同星珊瑚屬Plesiastrea Milne-Edwards & Haime
曲同星珊瑚 Plesiastrea curta (Dana)
多孔同星珊瑚 Plesiastrea versipora (Lamarck)
小星珊瑚屬Leptastrea
粗突小星珊瑚 Leptastrea bottae (Milne-Edwards & Haime)
刺星珊瑚屬Cyphastrea Milne-Edwards & Haime
鋸齒刺星珊瑚 Cyphastrea serailia (Forskal)
刺孔珊瑚屬Echinopora Lamarck
寶石刺孔珊瑚 Echinopora gemmacea ( Lamarck)
菊花珊瑚屬Goniastrea Milne-edwards & Haime
粗糙菊花珊瑚 Goniastrea aspera Verrill
濱珊瑚科Poritidae Gray, 1842
濱珊瑚屬Porites Link
澄黃濱珊瑚 Porites lutea Milne-Edwards & Haime
普哥濱珊瑚 Porites pukoensis Vaughan
扁縮濱珊瑚 Porites compressa Dana
角孔珊瑚屬Goniopora de Blainville
細角孔珊瑚 Goniopora gracilis (Bassett-Smith)
斯氏角孔珊瑚 Goniopora stutchburyi Wells
角孔珊瑚 Goniopora sp
裸肋珊瑚科Merulinidae Verrill, 1866
刺柄珊瑚屬 Hydnophora Fischer de Waldheim
鄰基刺柄珊瑚 Hydnophora contignatio (Forskal)
梳狀珊瑚科Pectiniidae Vaughan & Wells, 1943
尖孔珊瑚屬Oxypora Saville-Kent
撕裂尖孔珊瑚 Oxypora lacera (Verrill)
刺葉珊瑚屬Echinophyllia Klunzinger
粗糙刺葉珊瑚 Echinophyllia aspera (Ellis & Solander)
菌珊瑚科Agariciidae Gary, 1847
厚絲珊瑚屬Pachyseris Milne-Edwards & Haime
標準厚絲珊瑚 Pachyseris speciosa (Dana)
牡丹珊瑚屬Pavona Lamarck
十字牡丹珊瑚 Pavona decussata (Dana)
葉形牡丹珊瑚 Pavona frondifera Lamarck
易變牡丹珊瑚 Pavona varians Verrill
鐵星珊瑚科Siderastreidae Vaughan & Wells, 1943
假鐵星珊瑚屬Pseudosiderastrea Yabe & Sugiyama
假鐵星珊瑚 Pseudosiderastrea tayamai Yabe & Sugiyama
鹿角珊瑚科Acroporidae Verrill, 1902
薔薇珊瑚屬Montipora de Blainville
膨脹薔薇珊瑚 Montipora turgescens Bernard
鹿角珊瑚屬Acropora Oken
粗野鹿角珊瑚 Acropora humilis (Dana)
多孔鹿角珊瑚 Acropora millepora (Ehrenberg)
佳麗鹿角珊瑚 Acropora pulchra (Brook)
石松鹿角珊瑚 Acropora selago (Studer)
鹿角珊瑚 Acropora sp
枇杷珊瑚科Oculinidae Gray, 1847
盔形珊瑚屬Galaxea Oken
叢生盔形珊瑚 Galaxea fascicularis (Linnaeus)
稀杯盔形珊瑚 Galaxea astreata (Lamarck)
木珊瑚科Dendrophylliidae
筒星珊瑚屬Tubastraea
猩紅筒星珊瑚 Tubastrea coccinea Lesson
盾形陀螺珊瑚 Turbinaria peltata (Esper)
群棲錐形珊瑚 Balanophyllia generatrix cairn & Zibtowius
褶葉珊瑚科Mussidae Ortmann, 1890
葉狀珊瑚屬Lobophyllia de Blainville
傘房葉狀珊瑚 Lobophyllia corymbosa (Foskal)
軟珊瑚目Alcyonacea
軟珊瑚科Alcyonidae
短指軟珊瑚屬Sinularia
短指軟珊瑚 Sinularia sp
肉芝軟珊瑚屬Sarcophyton
肉芝軟珊瑚 Sarcophyton sp
豆莢軟珊瑚屬 Lobophytum
豆莢軟珊瑚 Lobophytum sp
棘軟珊瑚科 Nephtheidae
柔荑軟珊瑚屬 Nephthea
柔荑軟珊瑚 Nephthea sp
軟柳珊瑚科Subergorgiidae
軟柳珊瑚屬Subergorgia
側扁軟柳珊瑚 Subergorgia suberosa Pallas
粉灰軟柳珊瑚 Subergorgia ornate Thomson & Simpson
網狀軟柳珊瑚 Subergorgia reticulata(Ellis and Solander)
Subgorgia rubra
類尖柳珊瑚科 Paramuriceidae
小尖柳珊瑚屬 Muricella
扁小尖柳珊瑚 Muricella sibogae (Nutting)
中華小尖柳珊瑚 Muricella sinensis (Verrill)
小月柳珊瑚屬 Menella
長小月柳珊瑚 Menella praelonga (Ridley)
絨柳珊瑚屬 Villogorgia
緊絨柳珊瑚 Villogorgia compressa Hiles
Villagorgia antillarum
刺柳珊瑚屬 Echinogorgia
疏枝刺柳珊瑚 Echinogorgia pseudosassapo kollker
枝網刺柳珊瑚 Echinogorgia sassapo reticulate (Esper)
花刺柳珊瑚 Echinogorgia flora Nutting
珊瑚礁的位置楞刺柳珊瑚 Echinogorgia mertoni kunkenthal
鞭柳珊瑚科Ellisellidae
如燈芯柳珊瑚屬 Scirpearia
細如燈芯柳珊瑚 Scirpearia gracilis (Wright & Studer)
黃如燈芯柳珊瑚 Scirpearia erythraea Kukenthal
燈芯柳珊瑚屬Junceella
脆燈芯柳珊瑚 Junceella fragilis (Ridley)
燈芯柳珊瑚 Junceella juncea (Pallas)
鱗燈芯柳珊瑚 Junceella scuamata Toeplitz
蕾燈心柳珊瑚 Junceella gemmacea (Valenciennes)
叢柳珊瑚科 Plexauridae
厚叢柳珊瑚屬 Hicksonella
厚叢柳珊瑚 Hicksonella princeps Nutting
海底柏科Melithaidae
海底柏屬 Melithaea
赭色海底柏 Melithaea ocracea (Linnaeus)
疊葉柳珊瑚屬Mopsella
黃疊葉柳珊瑚 Mopsella rubeola (Wright & Studer)
群體海葵目ZOANTHIDEA
鞘群海葵科Epizoanthidae
皮群海葵屬Palythoa
皮群海葵 Palythoa sp.
基本分帶
 珊瑚礁
珊瑚礁珊瑚礁由3個基本相帶組成,即礁前帶、礁核帶和礁後帶:
①礁前帶。包括礁前塌積和礁前斜坡兩個亞相。前者位於後者的下方,坡度和緩,塌積物主要來源於礁核,粗細混雜,大礁塊直徑達數米。細小的生物碎屑也不少,這些沉積物在重力作用下,以崩塌和滑移等方式運移並堆積。礁前斜坡亦稱礁坪前坡,坡度達50°以上,其上限與珊瑚叢生帶呈過渡漸變關係,平面上呈槽溝與礁脊相間排列的鋸齒狀槽溝-礁脊系,礁前斜坡一般為硬底質。
②礁核帶。包括珊瑚叢生帶、礫堤(或藻脊)、礁坪和礁塘等4個亞相。珊瑚叢生帶的珊瑚不僅向海方增長,並以低潮線為上限向上繁衍,最終的結果是擴大礁坪。在波浪、潮流和風暴潮作用下,珊瑚叢生帶為礁體其他相帶提供了豐富的碳酸鹽沉積物。礫堤(或藻脊)展布在礁坪前緣,一般高出礁坪1米以上,抗浪性強,礫堤由礁塊或礁礫組成,物質來源於珊瑚叢生帶,通常由波浪、風暴潮上掀堆積。大洋環礁的礫堤,常被孔石藻、石葉藻等包殼粘生,形成藻脊。礁坪亦稱礁平台,是珊瑚礁的主體,主要由珊瑚礁岩構成。礁坪的發展上限為低潮線,表面因珊瑚群叢構成的圓桌狀突起(黑圓石)和不規則延伸的溶溝切割而崎嶇不平,有的還發育有珊瑚和貝殼碎屑組成的砂(或礫)斑堆積,礁坪上鬆散生物屑沉積物的粒徑由海向內變細。礁塘界於礁坪與海灘之間,深幾十厘米至數米,底質為砂質。
③礁後帶。包括海灘、砂壩(或灰砂島)和瀉湖3個亞相。海灘是向礁坪傾斜的潮間帶,由珊瑚屑等生物砂組成,岸礁的海灘有少量陸源碎屑,海灘砂可被文石、高鎂方解石等碳酸鹽類礦物膠結形成海灘岩;砂壩(或灰砂島)位於潮上帶,由珊瑚等生物砂(礫)組成,岸礁區的砂壩可見陸源碎屑,一般高3~5米;瀉湖是堡礁和環礁所特有的相,平均深度為45米,可分出瀉湖斜坡、點礁和湖底3個微相,瀉湖斜坡與點礁周邊有分枝狀珊瑚生長,湖底為灰泥(南海黃岩島環礁)或珊瑚泥(太平洋比基尼環礁)沉積。
上述相帶只有在堡礁和環礁才一應俱全;台礁無礁後帶;岸礁的礁後帶僅包括海灘和砂壩。
發展階段
 珊瑚礁
珊瑚礁1842年,達爾文提出的珊瑚礁成因的“沉降說”認為,珊瑚礁的形成經歷了三個發育階段:首先,在島嶼(尤其是火山島嶼)沿岸形成環繞海岸並與島嶼相連的岸礁;第二階段,島嶼略微下沉,珊瑚則與沉降速率保持同步向上生長,由於外緣海況條件好,珊瑚礁增長快於內側,結果珊瑚礁與海岸分開,中間隔以瀉湖,形成堡礁;第三階段,島嶼全部沉降入海,珊瑚仍向上生長,形成環繞瀉湖的環礁。這一理論的地質背景是火山島不斷下沉,所以又稱為“礁基沉降說”。達爾文還指出,某些環礁可在平坦的沉降台階上形成,不必經過堡礁階段;淺灘可直接由珊瑚生長發育成環礁,而無需發生沉降運動。沉降說將珊瑚礁三種主要類型密切聯繫成一完整的發生系列,獲得不少學者的支持。1950~1952年在馬紹爾群島埃尼威托克環礁鑽探,分別在1267米和1405米遇到玄武岩基底。由於珊瑚只能生長在淺水區,可見生長基底曾下沉1000餘米。對於大洋環礁來說,沉降說是可信的。
美籍加拿大學者R.A.戴利(1915)則強調冰川控制海平面變化,並對珊瑚礁發育產生影響,提出有關珊瑚礁成因的“冰川控制說”,認為冰期時海面下降100米,使出露的珊瑚死亡,當時水溫低,泥沙量大,珊瑚不能生存,已有的岸礁遭毀壞,灘地被削平到冰期時的海面高度。冰期後,海面上升100米,環境適合珊瑚繁衍,它們在冰期時形成的夷平台地上生長,一般是外緣更適合珊瑚繁殖,因而增長快,逐漸形成堡礁和環礁。戴利認為,冰川控制說能更好地說明瀉湖所具有的淺而平坦特性。但這一假說難以解釋鑽探揭露的厚逾千米的珊瑚礁體。
1935年,美國學者J.E.霍夫邁斯特和萊德提出關於環礁成因的“先成海台說”,但只能說明某些小而薄的礁體的成因。1947年,荷蘭地質學者P.H.奎年將沉降說和冰川控制說結合考慮,提出“冰期成因沉降說”,認為沉降說是正確的,但冰期海平面下降對珊瑚礁確有侵襲作用,對礁體的表面形態特徵有顯著影響。由於珊瑚礁的成因是複雜的,要圓滿解釋所有珊瑚礁的成因,還有待進一步努力。
研究意義
 珊瑚礁
珊瑚礁珊瑚從古生代初期開始繁衍,一直延續至今。珊瑚屬種多,演化快,常成為劃分地層的依據。造礁珊瑚對環境要求嚴格,只生長於熱帶、亞熱帶淺海中,而且隨著緯度升高,其屬種減少,生長率變慢,因而又可作為判斷古氣候、古地理的重要標誌。某些屬種的造礁珊瑚,每年會象樹木年輪那樣留下生長線,因而可將其當作“生物鐘”。寒武紀期間年生長線為424~412條,奧陶紀為412~402條,泥盆紀為410~385條,石炭紀為390條左右,三疊紀381條左右,現代為365條左右,這是地球自轉速度變慢的有力證據。
珊瑚礁與地殼運動有關。正常情況下,珊瑚礁形成於低潮線以下50米以淺的海域,高出海面者無疑是地殼上升或海平面下降的反映;反之,50米以深或覆蓋在平頂海山上的巨厚珊瑚礁灰岩,則標誌該處地殼下沉。根據珊瑚礁灰岩的產狀、厚度和分布等特點,還可以了解地殼運動的性質和特點。地槽區因地殼活動頻繁,升降幅度也大,常常會形成巨厚而結構複雜的珊瑚礁灰岩;地台區地殼較穩定,升降幅度不大,通常形成厚度不大、結構較簡單的珊瑚礁體;地台活化區由於地殼活動性大,個別地段甚至為較深的地塹和海灣,因而可形成較厚的珊瑚礁沉積。
地質時期珊瑚的生長北界不同,在晚侏羅世北界自英國經德國南部、阿爾卑斯山以北、高加索直到帕米爾一帶。以後,北界逐漸南移,到第四紀才與現代大體一致。這可能是氣候變遷和大陸漂移的結果。
珊瑚礁蘊藏著豐富的礦產資源。礁灰岩是多孔隙岩類,滲透性好,有機質豐度高,是油氣良好的生儲層。目前已發現和開採的礁型大油田有十多個,可采儲量50多億噸。礁型氣田也是高產的。大型油氣田多產於古代的堡礁中。珊瑚礁及其瀉湖沉積層中,還有煤炭、鋁土礦、錳礦、磷礦。礁體粗碎屑中發現銅、鉛、鋅等多金屬層控礦床。礁作為儲水層具有工業利用價值。珊瑚灰岩可作為燒制石灰、水泥的良好原料。有潮汐通道與外海溝通的環礁瀉湖,可闢為船舶的天然避風港。珊瑚礁灰岩覆蓋的平頂海山,可作為水下實驗的優良基地。千姿百態的珊瑚可作為裝飾工藝品。五彩繽紛的礁棲熱帶魚類可供人們觀賞。有些珊瑚早已被用作藥材。礁區具有豐富的漁業、水產資源。不少礁區已開闢為旅遊場所。
生物起源搖籃
珊瑚礁並非僅僅擁有美麗的外表以及棲息著豐富的物種。長期以來,它們一直充當著無數海洋生命的進化源泉的角色,其中甚至包括像蛤蜊和蝸牛這樣通常被科學家認為從淺海水域起源的物種。這一結論源自對化石記錄進行的新一輪調查,同時這一發現強化了進化潛能與環境具有重要關係的理論。
據美國《科學》雜誌線上新聞報導,珊瑚礁是眾所周知的生物多樣性熱點區域,但是科學家一直假設,許多棲息在珊瑚礁中的物種都是從其他生態系統中遷徙而來的,例如淺海水域。這項新研究的負責人、德國柏林自然史博物館的古生物學家Wolfgang Kiessling最初也認同這樣的假設。但是受到之前有關珊瑚礁研究的啟發,以及來自魚類遺傳學研究的提示,他又重新調查了這一問題。從2000年開始,Kiessling和兩名同事開始分析棲息在海底的動物的化石。在進行野外考察的基礎上,結合科學文獻,研究人員彙編了回溯至5.4億年前的全球各地的生物體記錄,其中包括大多數多細胞生物的進化歷史。Kiessling指出:“我們認為只有進行一次全面的研究才能夠回答我們提出的珊瑚礁是否是進化發源地的問題。”
根據化石第一次出現的位置,3位科學家確定了6615種海洋生物的起源環境。他們在1月8日出版的美國《科學》雜誌上報告說,大約有1426種生物起源於珊瑚礁中,這幾乎比淺水環境中的生物起源數量多了50%。此外,Kiessling表示,他們發現,珊瑚礁對其他棲息地的生物多樣性亦作出了貢獻,這是因為一些起源於珊瑚礁生物系統的物種後來又遷徙到了別處。他說:“我們對於珊瑚礁作為生物起源搖籃的作用是如此巨大感到非常吃驚。”
美國墨爾本市佛羅里達理工學院的古生物學家Richard Aronson表示:“這是一篇非常吸引人並且極為重要的論文。”他說:“這意味著如果現代珊瑚礁持續遭到破壞,那么將因為切斷新的生物多樣性的供給而使其他生態系統的進化遭受長期影響。”
威脅
對於珊瑚礁來說人類是唯一的巨大威脅。尤其陸地上的污染和過渡捕撈對這些生態系統造成了嚴重威脅。船隻了拖網造成的物理破壞也是一個問題。活魚貿易導致了使用少量氰化物和其他化學藥劑來捕捉小魚的手段。此外氣候現象如厄爾尼諾現象和全球變暖造成的過高的水溫也會導致珊瑚白化。按照大自然保護協會的數據目前全球珊瑚礁破壞速度不斷加快,在50年內全球70%的珊瑚礁會消失。對於生活在熱帶的人來說這個損失可能意味著一個大災難。2003年修等寫道:“隨著人口的增長,以及運輸和儲存系統的發展,人類對珊瑚礁的影響的發展呈指數倍增長。比如對魚和其他自然資源的市場需求全球化,對珊瑚礁資源的需求比對熱帶資源的需求的增長快得多。”
目前學者還在研究各種因素對珊瑚礁系統的影響。這些因素的列表很長,從海洋吸收二氧化碳開始,到大氣層的變化、紫外線的影響、海洋酸化、病毒、沙暴將病菌帶到遠海珊瑚礁的可能性、不同的污染物等等。不但近海的珊瑚礁受到威脅,因此不僅陸上的發展和污染是一個問題。
陸上發展和污染
土地使用過度或者管理不良均會威脅珊瑚礁。在過去20年中,過去大量在海岸出現的紅樹林由於造路、建築工程、機場海港建築、隧道建築等被破壞。這些紅樹林能夠吸收許多從陸地上流失的營養。從陸地上流失的營養在海水中會導致藻類和浮游生物的大量繁殖,形成赤潮。而珊瑚礁則需要營養少的海水,需要充分的陽光。大量的營養流入會破壞其生態系統內部的平衡。海岸濕地的喪失和赤潮是嚴重影響珊瑚礁所需要的水質的因素。
水質差的水似乎也促進珊瑚蟲傳染病的傳播。
工業污染物中很常見的銅影響珊瑚蟲的生長。
活魚貿易
由於北美和歐洲對珊瑚礁觀賞魚的需求在印度洋-太平洋地區使用氰化物捕魚的方法大增。世界上85%的觀賞魚是在這個地區捕取的,幾乎所有這些魚是使用氰化物捕取的。捕魚人使用氰化物來麻醉魚,然後可以輕易地捕取它們。使用這個手段捕魚魚的死亡率達90%。氰化物對珊瑚礁生態系統的破壞也很大,它殺死珊瑚蟲和其他無脊椎動物。同時通過捕走對整個生態系統平衡必須的魚本身就對珊瑚礁造成了威脅。有時捕魚人還敲擊珊瑚礁來驚嚇魚,讓它們從縫隙中逃出來,或者破壞珊瑚礁來捕取藏在縫隙中的魚。
貧困是促使氰化物捕魚普及的重要原因。有些地區如菲律賓使用氰化物捕捉活魚非常普及而且是合法的,而當地40%的人生活在貧困線以下。在這些開發中國家中捕魚人只有依靠這樣的手段才能養活自己和家人。
另一種對珊瑚礁破壞巨大的捕魚方法是使用炸藥捕魚。捕魚的人使用一個裝有硝酸鉀的瓶子。瓶子爆炸時在水下造成一個衝擊波,導致魚鰾破裂。之後魚浮到水面上。捕魚的人往往在第一次爆炸後再進行第二次爆炸來捕取被死魚吸引來的食肉魚。這個手段不但炸死許多小魚而且還炸死許多生活在珊瑚礁里的其他生物,甚至於珊瑚礁本身。過去充滿珊瑚的地方今天變成一片荒漠,沒有一點珊瑚,就更不用說其他生活在珊瑚礁中的生物了。
 白化的珊瑚礁
白化的珊瑚礁珊瑚白化
在1998年和2004年的厄爾尼諾現象過程中海水水溫超過了一般情況,許多珊瑚礁出現了白化的現象,有些死亡。此後部分離污染源遠的地區的珊瑚礁獲得恢復。但是也有些學者認為由於全球變暖造成的珊瑚礁的擴展會高於其死亡。有人甚至估計到2100年全球珊瑚礁的面積會比工業革命前增長35%。
關於珊瑚白化的原因以及全球變暖的影響現在在學術界還有很大的爭議。
全球的破壞情況
東南亞的珊瑚礁主要受到捕魚(使用氰化物和炸藥)、過量捕撈、泥沙沉積、污染和白化的威脅。通過教育、管理和設立海洋保護區的方式各個國家試圖保護這些珊瑚礁。比如印度尼西亞擁有3.3萬平方海里的珊瑚礁,占全世界的1/3,印度尼西亞1/4的魚類生活在這些珊瑚礁內。這些珊瑚礁受到破壞性捕魚、無管理的旅遊業和由於氣候變化導致的白化的破壞。2000年印度尼西亞414個珊瑚礁觀察站提供的數據表明印尼僅6%的珊瑚礁處於完好的狀態,24%處於良好狀態,約70%處於惡劣至中等狀態。
粗的估計全球約10%的珊瑚礁近乎死亡。威脅原因如上所述從捕魚技術的環境影響直到海洋的酸化。珊瑚白化也是一個全球性的問題。
保護和重建
估計全球60%的珊瑚礁受到人類活動所造成的威脅。尤其在東南亞威脅特別嚴重,在這裡80%的珊瑚礁處於危險狀態。
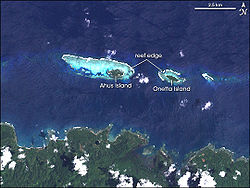 巴布亞紐幾內亞馬努斯省的居民出於傳統原因禁止一定的捕魚方法,因此這裡的珊瑚礁的總生物質能和魚的大小均比相應其他地區的高
巴布亞紐幾內亞馬努斯省的居民出於傳統原因禁止一定的捕魚方法,因此這裡的珊瑚礁的總生物質能和魚的大小均比相應其他地區的高海事保護區
一個越來越廣泛被採納的措施是設立海事保護區。在東南亞和全球其他地區均有引入海事保護區來改善漁業管理和保護生態環境的試圖。這些保護區相當於陸地上的自然公園或者野生保護區。在這裡不允許捕魚。設立海事保護區即有生態目的,也有社會目的。它旨在使得珊瑚礁得到恢復、維持其美觀、增加和保護其生物多樣性,以及改善當地人的經濟狀況。不落實、相互矛盾的見解和財政來源是設立保護區最大的困難。
目前印度尼西亞有九個海事保護區,總面積41,129平方公里。一個在一個最近建立的保護區進行的研究發現要成功地管理這樣的一個保護區需要國家、省份和當地居民的共同合作。
保護組織
許多組織的目的是幫助保護珊瑚礁。比如珊瑚礁聯盟是一個由其成員支持的、非營利組織。這個組織通過綜合性的生態系統管理、可持續旅遊業以及社區合作等手段來保護珊瑚礁的健康。它與當地人合作來分析和解決保護問題、通過教育和訓練來改變當地人的看法、提供資源來加強保護的努力、產生可持續旅遊業的刺激等等。它目前的活動範圍包括佛羅里達、貝里斯、宏都拉斯、墨西哥、斐濟、巴布亞紐幾內亞和夏威夷。
珊瑚礁缸中加鈣的方法
天然海水中,鈣離子濃度為每公斤400毫克。珊瑚礁四周或沿岸附近海水的鈣離子濃度略微高一些約為420毫克-480毫克/升。由此推知無脊椎動物生態缸的鈣離子濃度應在400毫克-500毫克/升之間。如果你對無脊椎動物生態缸沒有添加過鈣質,那么鈣離子可能就只有250毫克-350毫克/升。此時您要注意添加鈣離子,有兩種方法添加鈣。氯化鈣能在水中迅速地大量溶解而且加入水族箱內不會影響pH值。但它也有兩個缺點,一是過一些時間後,水族箱會產生鈉離子對氯離子比率不平衡的結果。另一個缺點是氯化鈣過量會導至鈣離子濃度快速上升,可能會引起鹼度降低,其後需碳酸鹽緩衝系統加以平衡。建議你用鈣離子濃度超過400毫克/升的石灰水調配入工海水,每周加一點石灰水,並常測PH值的變化。你會發現粉紅色和紫色外皮的珊瑚共生藻會擴散開來覆蓋珊瑚的赤裸表面,珊瑚會繼續生長,生態缸漂亮多了。將氫氧化鈣或氧化鈣溶在水中,輕輕地混合溶液並放置數小葉直到石灰水清澈為止,白色未溶解的氫氧化鈣會沉澱在底部,用缸吸管將清澈的石灰水吸取出來,剩下的未溶解部分倒掉。注意:你在添加石灰水時,不要一次加太多,因為石灰水的PH值幾乎高達12,即使添加少許石灰水,也會暫時生使生態缸的PH值升高,如果一下子添加多了會使pH值急劇上升而傷害魚類,甚至死亡。所以要慢慢地加入石灰水。
海洋學相關知識(六)
| 海洋科學是研究海洋的自然現象、性質及其變化規律,以及與開發利用海洋有關的知識體系。下面讓我們來完善與海洋學相關的詞條,以此來更加深入的了解海洋學的相關知識。 |
