
概述
免疫學和遺傳學交叉的邊緣學科,主要研究免疫系統的結構和功能如免疫應答、抗體的多樣性等的遺傳基礎。此外,也套用免疫學的方法來識別個體間的遺傳差異(如血型、表面抗原等)以作為遺傳規律分析的指標。免疫遺傳學是現代醫學臨床實踐的重要理論基礎之一,是輸血、器官移植、胎母不相容和親子鑑定的理論基礎,對闡明免疫系統的演化、人種差異和生物進化也有重要意義。
簡史
1900~1902年奧地利醫學家K.蘭德施泰納等交叉混和不同人的血清和紅細胞,通過細胞凝集反應的觀察首次檢出人的兩種紅細胞同種異體抗原A和B。1924年F.伯恩斯坦證明 ABO血型分別為三個復等位基因所控制,這是第一個被發現的人的血型遺傳系統。此後在一些動物和人體中陸續發現了多種紅細胞抗原(見血型遺傳),並逐步闡明了它們的遺傳規律。1958年法國免疫學家J.多塞發現了人的第一個白細胞抗原。1959年澳大利亞免疫學家F.M.伯內特提出克隆選擇學說,從細胞水平說明了抗體多樣性的來源。70年代分子遺傳學和體細胞遺傳學的發展進一步推動了免疫遺傳學的發展。例如套用遺傳工程和分子雜交技術證明了抗體分子的重鏈和輕鏈各由若干DNA序列所分別編碼;套用體細胞雜交創建了製備單克隆抗體的技術。這些新的發展使免疫遺傳學的研究成為分子遺傳學研究的重要內容。
研究內容
免疫遺傳學的內容主要包括抗原遺傳、抗體遺傳和免疫應答遺傳三個方面。
抗原遺傳
①紅細胞抗原(見血型遺傳)。②白細胞抗原,1936年P.A.戈勒用兔抗小鼠紅細胞血清檢出小鼠的四種紅細胞抗原。他使抗原Ⅱ陽性小鼠品系與抗原Ⅱ陰性小鼠品系雜交,再使子代與抗原Ⅱ陰性的小鼠品系回交,回交子代或是抗原Ⅱ陽性或是陰性。將抗原Ⅱ陽性小鼠的腫瘤移植給抗原Ⅱ陰性子代則腫瘤被排斥;移植給抗原Ⅱ陽性子代則腫瘤不被排斥,證明了抗原Ⅱ是一種組織相容性抗原(H-2),由單一基因所控制。接著又發現H-2特異性可以在淋巴細胞上檢出,開創了白細胞抗原的研究。以後證明小鼠的組織相容性抗原並不是由單個基因而是由若干緊密連鎖的基因所控制。這許多基因構成主要組織相容性複合體(MHC),它位置在小鼠第17號染色體上,分為K、I、S、G、D/L、T6個區。人體的主要組織相容性複合體稱為人白細胞抗原,簡稱 HLA。決定HLA的複合座位在 6號染色體短臂上,已知有A、C、B、D/DR幾個座位,每個座位都有很多共顯性復等位基因。猴和狗的相應的基因複合體分別稱為 RhLA和DLA。③其他動物的抗原遺傳,各種動物細胞的蛋白質都是抗原物質,這些蛋白質的差異都可用免疫學方法檢出,從而揭示其遺傳機理。例如套用兔抗草履蟲碎片的抗血清,發現每種草履蟲品系可合成大約10種不同特異性的表面抗原。雙小核草履蟲有s、g、d3個主要基因控制它的表面抗原S、G、D。這些基因都不連鎖。這些表面抗原雖受核基因控制,但是它們的表達受溫度的影響。在15~18℃中表達s基因,隨著溫度升高s基因關閉而表達g基因,溫度大約30~32℃則表達 d基因。不同地理分布的不同品系例如156和168品系的 g基因可以編碼不同的抗原156G和168G,但當這些不同品系的個體經接合生殖形成雜合體後,雖然它們的基因型相同(都是156g/168g),培養時的溫度條件也相同(30℃),但原來屬於156品系的雜合體草履蟲的無性繁殖系則表達抗原G,而原來屬於168品系的雜合體草履蟲的無性繁殖系則表達抗原 D。這種基因型相同的雜合體在一定情況下出現不同的表型,並且可以通過無性繁殖世代相傳的現象稱為後成遺傳現象。後成遺傳的本質既是遺傳學也是免疫學中的一個沒有得到完滿解答的問題。
抗體遺傳
抗體分子是免疫球蛋白,由兩條輕鏈和兩條重鏈組成。輕鏈和重鏈都可按胺基酸順序變異的程度劃分為可變區(V)和恆定區(C)。高等動物和人體能夠產生極其大量的不同特異性的免疫球蛋白。是不是每個個體都有這么多的輕鏈基因和重鏈基因?這曾使遺傳學家迷惑不解。伯內特的克隆選擇學說在細胞水平上回答了這一問題。按照這一學說,每一個漿細胞只能產生一種或少數幾種抗體,一個個體的無數的漿細胞合在一起就可產生出無數種類的抗體分子。
多種多樣的漿細胞是由淋巴細胞分化而來的。套用基因建株、分子雜交和核苷酸順序分析技術,發現免疫球蛋白分子的輕鏈和重鏈都是由若干被隔開的基因片段或稱外顯子編碼的。在決定輕鏈的染色體上,有L、V、J、C四類基因片段。V片段(可變片段)約有150種,J片段(連線片段)約有5種,L片段(引導片段)和C片段(恆定片段)各有一種。在淋巴細胞分化過程中,這些基因片段經過重排而連線在一起後才能被轉錄。重排時 V/J接頭有靈活性,估計有10種可能性。原始轉錄產物再經過加工拼接,才成為L-V-J-C連續的mRNA。L片段在翻譯後被切去,V-J片段翻譯成為可變區,C片段翻譯成為恆定區。重排時的不同組合和 V/J接頭的靈活性可導致約7500種(150×5×10)輕鏈基因(圖1)。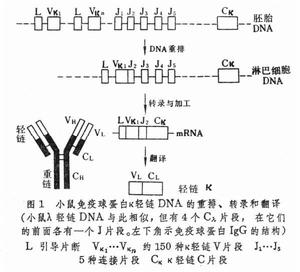 在決定重鏈子的染色體上,有L、V、D、J、C五類基因片段。其中L片段1種,估計V片段有80種,D片段(多樣性片段)有50種,J片段有6種,C片段有 8種。V/D和D/J接頭估計各有10種可能性。8種C片段決定免疫球蛋白的類別,即lgM、lgD、lgG3、lgG1、lgG2b、lgG2a、lgE或lgA,它們針對同一種抗原決定簇而有同樣的可變區。通過重排和接頭的靈活性可導致240萬種(80×50×6×10×10)重鏈基因(圖2)。輕鏈和重鏈組合起來可以產生180億種(750×2400000)免疫球蛋白分子。如果再加上可能產生的體細胞突變(估計突變率是1/10000細胞/代),抗體的多樣性還可以擴大。抗體多樣性的來源雖然已經在細胞水平和分子水平上得到了一些說明,但是還有許多問題有待解決。已知一種漿細胞只產生一種特異性抗體分子,在雜合狀態下它的等位基因是不表達的。對這種等位排斥現象至今不知道它的機理。對於染色體重排的確切過程現在知道得也還很少。
在決定重鏈子的染色體上,有L、V、D、J、C五類基因片段。其中L片段1種,估計V片段有80種,D片段(多樣性片段)有50種,J片段有6種,C片段有 8種。V/D和D/J接頭估計各有10種可能性。8種C片段決定免疫球蛋白的類別,即lgM、lgD、lgG3、lgG1、lgG2b、lgG2a、lgE或lgA,它們針對同一種抗原決定簇而有同樣的可變區。通過重排和接頭的靈活性可導致240萬種(80×50×6×10×10)重鏈基因(圖2)。輕鏈和重鏈組合起來可以產生180億種(750×2400000)免疫球蛋白分子。如果再加上可能產生的體細胞突變(估計突變率是1/10000細胞/代),抗體的多樣性還可以擴大。抗體多樣性的來源雖然已經在細胞水平和分子水平上得到了一些說明,但是還有許多問題有待解決。已知一種漿細胞只產生一種特異性抗體分子,在雜合狀態下它的等位基因是不表達的。對這種等位排斥現象至今不知道它的機理。對於染色體重排的確切過程現在知道得也還很少。
免疫應答遺傳
早在1938年P.A.戈勒等就發現小鼠不同近交系對肺炎球菌莢膜多糖產生抗體的能力大不相同。後來有人以人工合成的多肽作為抗原,也得到同樣的結果。這種差別是遺傳的。免疫應答基因(Ir)控制一個個體對抗原發生免疫應答的程度。例如以一種人工合成的多肽免疫小鼠,H-2系統q純合小鼠(Irq/Irq)是高應答品系,能產生較多的抗體;s純合小鼠(Irs/Irs)是低應答品系,產生的抗體較少。它們的雜交子代(Irq/Irs)與q純合親代相同,也是高應答的。將雜交子代與s純系親代回交,回交子代半數是高應答的,半數是低應答的。這一事實說明免疫應答反應由一對基因所控制,高應答反應是顯性,低應答反應是隱性。小鼠有幾個Ir基因,有時對一種抗原的免疫應答可同時受兩個Ir基因控制。人體對豕草變應原產生lgE抗體的應答程度也受與HLA連鎖的Ir基因控制。在雞、大鼠、狗、猴等動物中也都發現有Ir基因。
除上述三個方面的內容外,補體的遺傳和干擾素的遺傳也屬於免疫遺傳學研究的範疇。
免疫系統
| 系統 | 自適應與天然 | 體液與細胞 | 補體(過敏毒素) | 固有 |
| 抗原與抗體 | 抗原 (超抗原;變應原) | 半抗原 |
| 表位 (線性;構象) | |
| 抗體 (單克隆抗體; 多克隆抗體;自身抗體) | 多克隆B細胞反應 | 同種異型 | 同種型 | 遺傳型 | |
| 免疫複合物 | |
| 免疫細胞/白細胞 | 淋巴系細胞: T細胞 | B細胞 | NK細胞 |
| 髓系細胞:肥大細胞 | 嗜鹼性粒細胞 | 嗜酸性粒細胞 | 巨噬細胞 | |
| 吞噬細胞:中性粒細胞 | 巨噬細胞/網狀內皮系統 | |
| 專職型 APCs:樹突狀細胞 | 巨噬細胞 | B細胞 | |
| 免疫與耐受 | 作用:免疫性 | 自身免疫 | 變態反應 | 炎症 | 交叉反應性 |
| 無作用:耐受 (中樞耐受;外周耐受;克隆無能;克隆缺失) | 免疫缺損 | |
| 免疫遺傳學 | 體細胞超突變 | V(D)J 重組 | 免疫球蛋白類別轉換 | MHC/HLA |
| 免疫物質 | 細胞因子 | 調理素 | 溶細胞素 |
| 其他 | 診斷免疫學 |
免疫系統/ 免疫學
| 免疫系統 | 它是人體抵禦病原菌侵犯最重要的保衛系統。 |
| 系統 | |
| 抗原與抗體 | 抗原 (超抗原, 變應原) | 半抗原 表位 (線性表位, 構象表位) 抗體 (單克隆抗體, 多克隆抗體, 自身抗體) | 多克隆B細胞反應 | 同種異型 | 免疫球蛋白同種異型 | 遺傳型 | 免疫複合物 |
| 免疫細胞/白細胞 | 淋巴系細胞:T細胞 | B細胞 | NK細胞 髓系細胞:肥大細胞 | 嗜鹼性粒細胞 | 嗜酸性粒細胞 | 巨噬細胞 吞噬細胞:中性粒細胞 | 巨噬細胞 | 網狀內皮系統 抗原遞呈細胞:樹突狀細胞 | 巨噬細胞 | B細胞 |
| 免疫 與 耐受 | 作用:免疫性 | 自身免疫 | 變態反應 | 炎症 | 交叉反應性 無作用:耐受 (中樞耐受, 外周耐受, 克隆無能, 克隆缺失) | 免疫缺損 |
| 免疫遺傳學 | 體細胞超突變 | V(D)J 重組 | 免疫球蛋白類別轉換 | MHC/HLA |
| 免疫物質 | 細胞因子 | 調理素 | 溶細胞素 |
| 其他 | 診斷免疫學 |
