
 重組DNA技術
重組DNA技術概述
基因工程是在分子水平上對基因進行操作的複雜技術,一般包括4個步驟:一是克隆目的基因,取得所需要的·DNA特異片段;二是將目的基因與DNA載體連線成重組DNA;三是將重組DNA引入細菌或動植物細胞內使其增殖;四是將表達目的基因的受體細胞挑選出來,使目的基因表達相應的蛋白質或其他產物,從而育成動植物優良新品種(系)。自1977年成功地用大腸桿菌生產出生長Itl釋放抑制因子以來,人胰島素、人生長激素、胸腺肽、干擾素、尿激酶、腫瘤壞死因子、瘋牛病疫苗、B型肝炎病毒疫苗、A型肝炎病毒疫苗、幼畜腹瀉疫苗和青黴素醯化酶基因工程菌株等數十種基因工程產品相繼問世。優質產毛羊等動物新品種,金色水稻,抗蟲或抗除草劑的玉米、大豆、棉花、水稻,轉類胡蘿蔔素生物合成相關酶基因花卉、蔬菜等已獲推廣或已取得階段性成果。
基本簡介
 重組DNA技術
重組DNA技術重組DNA技術來源於兩個方面的基礎理論研究——限制性核酸內切酶(簡稱限制酶)和基因載體(簡稱載體)。限制酶的研究可以追溯到1952年美國分子遺傳學家S.E.盧里亞在大腸桿菌中所發現的一種所謂限制現象——從菌株甲的細菌所釋放的噬菌體能有效地感染同一菌株的細菌,可是不能有效地感染菌株乙;少數被感染的菌株乙的細菌所釋放的同一噬菌體能有效地感染菌株乙可是不能有效地感染菌株甲。經過長期的研究,美國學者W.阿爾伯在1974年終於對這一現象提出了解釋,認為通過噬菌體感染而進入細菌細胞的DNA分子能被細菌識別而分解,細菌本身的DNA則由於已被自己所修飾(甲基化)而免於被分解。但有少數噬菌體在沒有被分解以前已被修飾了,這些噬菌體經釋放後便能有效地感染同一菌株的細菌。被甲(或乙)這一菌株所修飾的噬菌體只能有效地感染甲(或乙),而不能有效地感染乙(或甲),說明各個菌株對於外來DNA的限制作用常常是專一性的。通過進一步的研究發現這種限制現象是由於細菌細胞中具有專一性的限制性核酸內切酶的緣故。
重組DNA技術中所用的載體主要是質粒和溫和噬菌體(見轉導)兩類,而在實際套用中的載體幾乎都是經過改造的質粒或溫和噬菌體。英國微生物遺傳學家W.海斯和美國微生物遺傳學家J.萊德伯格等在1952年首先認識到大腸桿菌的F因子(見細菌接合)是染色體外的遺傳因子。1953年法國學者P.弗雷德里克等發現大腸桿菌產生大腸桿菌素這一性狀為一種染色體外的大腸桿菌素因子所控制。1957年日本學者發現了抗藥性質粒。後兩類質粒都是在遺傳工程中廣泛套用的質粒。
重組DNA技術中廣泛套用的噬菌體是大腸桿菌的溫和噬菌體λ,它是在1951年由美國學者E.萊德伯格等發現的。到70年代初,生物化學研究的進展也為重組DNA技術奠定了基??972年美國的分子生物學家P.伯格等將動物病毒SV40的DNA與噬菌體P22的DNA連線在一起,構成了第一批重組體DNA分子。1973年美國的分子生物學家S.N.科恩等又將幾種不同的外源DNA插入質粒pSC101的DNA中,並進一步將它們引入大腸桿菌中,從而開創了遺傳工程的研究。
相關概念
·克隆與克隆化
 重組DNA技術表格介紹
重組DNA技術表格介紹·DNA克隆
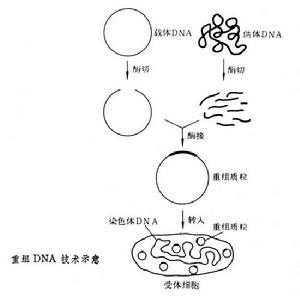
DNA克隆就是套用酶學的方法,在體外將各種來源的遺傳物質與載體DNA結合成一具有自我複製能力的DNA分子——複製子,繼而通過轉化或轉染宿主細胞、篩選出含有目的基因轉化子細胞,再進行擴增、提取獲得大量同一DNA分子,即DNA克隆又稱重組DNA。
·工具酶
在重組DNA技術中,常需要一些基本工具酶進行基因操作。小結:重組DNA技術常用工具酶(1)限制性內切酶:識別特異序列,切割DNA。(2)DNA連線酶:催化DNA中相鄰的5′磷酸基與3′羥基間形成磷酸二酯鍵,使DNA切口封合,連線DNA片段。(3)DNA聚合酶Ⅰ:a.合成雙鏈cDNA中第二條鏈。b.缺口平移製做探針。c.DNA序列分析。d.填補3′末端。(4)Taq酶催化PCR反應,聚合DNA。(5)反轉錄酶a.合成cDNA。b.替代DNA聚合酶Ⅰ進行填補,標記或DNA序列分析,(6)多聚核苷酸激酶催化DNA5′羥基末端磷酸化,或標記探針。(7)鹼性磷酸酶切除DNA5′末端磷酸基。(8)末端轉移酶在3′羥基末端進行同系多聚核苷酸加尾。(9)DNA酶:切割DNA(10)RNA酶:切割RNA。
在所在工具酶中,限制性核酶內切酶具有特別重要的意義。所謂限制性核酸內切酶就是識別DNA的特異序列,並在識別點或其周圍切割雙鏈DNA的一類內切酶。根據酶的組成,所需因子及裂解DNA方式的不同,可將限制性核酸內切酶分為三類。重組DNA技術中常用的限制性核酸內切酶為Ⅱ類酶,大部分Ⅱ類酶識別DNA位點的核苷酸序列呈二元鏇轉對稱,通常稱這種特殊的結構順序為迴文結構。
步驟和技術路線
 重組DNA技術技術路線
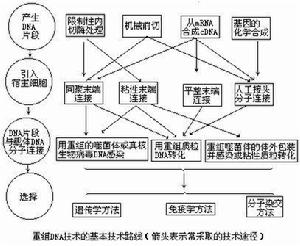
重組DNA技術技術路線重組DNA片段的取得主要的方法有:①利用限制酶取得具有粘性末端或平整末端的DNA片段;②用機械方法剪下取得具有平整末端的DNA片段,例如用超音波斷裂雙鏈DNA分子;③經反向轉錄酶的作用從mRNA獲得與mRNA順序互補的DNA單鏈,然後再複製形成雙鏈DNA(cDNA)。例如人的胰島素和血紅蛋白的結構基因都用這方法獲得。這樣獲得的基因具有編碼蛋白質的全部核苷酸順序,但往往與原來位置在染色體上的基因在結構上有區別,它們不含有稱為內含子的不編碼蛋白質的間隔順序(見基因);④用化學方法合成DNA片段。從蛋白質肽鏈的胺基酸順序可以知道它的遺傳密碼。依照這密碼用化學方法可以人工合成基因。
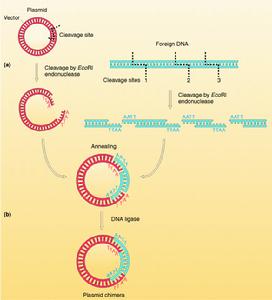
DNA片段和載體的連線DNA片段和載體相連線的方法主要有四種:①粘性末端連線,每一種限制性核酸內切酶作用於DNA分子上的特定的識別順序,許多酶作用的結果產生具有粘性末端的兩個DNA片段。例如來自大腸桿菌(Escherichiacoli)的限制酶EcoRI作用於識別順序↓…GAATTC……CTTAAG…↑(↑指示切點),產生具有粘性末端…G…CTTAA和AATTC…G…的片段。把所要克隆的DNA和…載體DNA用同一種限制酶處理後再經DNA連線酶處理,就可以把它們連線起來。②平整末端連線,某些限制性內切酶作用的結果產生不含粘性末端的平整末端。例如來自副流感嗜血桿菌(Hemophilusparainfluenzae)的限制酶Hpal作用於識別順序↓…GTTAAC……CAATTG…而產生末端為…GTT…GAA的DNA片段。用機械剪下方法取得的DNA片段的末端也是平整的。在某些連線酶(例如感染噬菌體T4後的大腸桿菌所產生的DNA連線酶)的作用下同樣可以把兩個這樣的DNA片段連線起來。③同聚末端連線,在脫氧核苷酸轉移酶(也稱末端轉移酶)的作用下可以在DNA的3′羧基端合成低聚多核苷酸。如果把所需要的DNA片段接上低聚腺嘌呤核苷酸,而把載體分子接上低聚胸腺嘧啶核苷酸,那么由於兩者之間能形成互補氫鍵,同樣可以通過DNA連線酶的作用而完成DNA片段和載體間的連線。④人工接頭分子連線,在兩個平整末端DNA片段的一端接上用人工合成的寡聚核苷酸接頭片段,這裡面包含有某一限制酶的識別位點。經這一限制酶處理便可以得到具有粘性末端的兩個DNA片段,進一步便可以用DNA連線酶把這樣兩個DNA分子連線起來。
導入宿主細胞將連線有所需要的DNA的載體導入宿主細胞的常用方法有四種:①轉化,用質粒作載體所常用的方法。②轉染(見轉化),用噬菌體DNA作載體所用的方法,這裡所用的噬菌體DNA並沒有包上它的外殼。③轉導,用噬菌體作載體所用的方法,這裡所用的噬菌體DNA被包上了它的外殼,不過這外殼並不是在噬菌體感染過程中包上,而是在離體情況下包上的,所以稱為離體包裝。④注射,如果宿主是比較大的動植物細胞則可以用注射方法把重組DNA分子導入。
選擇用以上任何一種方法連線起來的DNA中既可能包括所需要的DNA片段,也可能包括並不需要的片段,甚至包括互相連線起來的載體分子的聚合體。所以接受這些DNA的宿主細胞中間只有一小部分是真正含有所需要的基因的。一般通過3種方法可以取得所需要的宿主細胞:①遺傳學方法,對於帶有抗藥性基因的質粒來講,從被轉化細菌是否由敏感狀態變為抗藥的狀態就可以知道它有沒有獲得這一抗藥性質粒。一個抗藥性基因中間如果接上了一段外來的DNA片段,就使獲得這一質粒的細菌不再表現抗性。把一個帶有兩個抗性基因氨苄青黴素抗性和四環素抗性的質粒pBR322用限制酶BamHI處理,由於BamHI的唯一的識別位點是在四環素抗性基因中,所以經同一種酶處理的DNA分子片段就可以連線在這一基因中間。在被轉化的細菌中選擇只對氨苄青黴素具有抗性而對四環素不具抗性的細菌,便可以獲得帶有外來DNA片段的載體的細菌。這是一種常用的遺傳學方法。②免疫學方法和分子雜交方法,當一個宿主細胞獲得了攜帶在載體上的基因後,細胞中往往就出現這一基因所編碼的蛋白質,用免疫學方法可以檢出這種細胞。分子雜交的原理和方法同樣可以用來檢測這一基因的存在(見分子雜交、基因文庫)。
基因表達在構建重組體DNA分子和選擇宿主細胞時,還須考慮外源基因表達的問題。就是說要求外來的基因在宿主細胞中能準確地轉錄和翻譯,所產生的蛋白質在宿主細胞中不被分解,而且最好還能分泌到細胞外。為了使外源基因表達,需要在基因編碼順序的5′端有能被宿主細胞識別的啟動基因順序以及核糖體的結合順序。兩種常用的方法能用來使外源基因在宿主細胞中順利地表達:①在形成重組體DNA分子時在載體的啟動基因順序和核糖體結合順序後面的適當位置上連線外源基因。例如將兔的β-珠蛋白基因或人的成纖維細胞干擾素基因分別連線到已經處在載體上的大腸桿菌乳糖操縱子的啟動基因後面,便能使它們在大腸桿菌中順利地表達;②將外源基因插入到載體的結構基因中的適當位置上,轉錄和翻譯的結果將產生一個融合蛋白。這種融合蛋白質被提純後,還要準確地將兩部分分開,才能獲得所需要的蛋白質。在早期的遺傳工程研究中,生長激素釋放抑制因子和鼠胰島素基因的表達都是通過將它們連線在β-半乳糖苷酶基因中的方式實現的。
套用介紹
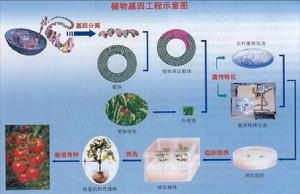 重組DNA技術
重組DNA技術利用遺傳工程手段還可以提高微生物本身所產生的酶的產量。例如可以把大腸桿菌連線酶的產量提高500倍。
理論研究套用重組DNA技術可以克隆和擴增某些原核生物和真核生物的基因,從而可以進一步研究它們的結構和功能。重組DNA技術的成就和提出的問題促進了遺傳學、生物化學、微生物學、生物物理學和細胞學等學科的發展,並且有助於這些不同學科的結合。目前正在形成一門新興的學科——生物工藝學或生物工程學,就是這種趨勢的反映。
動植物育種和基因治療已經有一些研究工作明確地預示著重組DNA技術在這些方面的潛力。例如把來自兔的β-血紅蛋白基因注射到小鼠受精卵的核內,再將這種受精卵放回到小鼠輸卵管內使它發育,在生下來的小鼠的肝細胞中發現有兔的β-血紅蛋白基因和兔的β-血紅蛋白。還有人把包括小鼠的金屬巰基組氨酸三甲基內鹽I(metallothioneineI)基因的啟動子及大鼠生長激素結構基因的DNA片段注射進小鼠受精卵的前核中,由此發育得來的一部分小鼠由於帶有可表達的大鼠生長激素基因,所以明顯地比對照鼠長得大。這些實驗結果為基因治療展現了可喜的前景。固氮的功能涉及17個基因,分屬7個操縱子,現在已能把它們全部引入酵母菌,而且能正常地複製,不過還沒有能使這些基因表達。改造玉米胚乳蛋白質而使人畜營養必需的賴氨酸和色氨酸成分增加的工作也在著手進行。大豆的基因已能通過Ti質粒引入向日葵。因此,可以預期隨著時間的推移在能源、農業、食品生產、工業化學和藥品製造等方面都將會取得巨大的成果。
內容評價
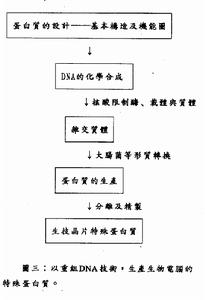 重組DNA技術
重組DNA技術重組DNA技術或遺傳工程最初是用來將DNA片段克隆到微生物宿主中,以過表達特定的基因產物用於進一步研究。重組DNA分子也已經用於獲得遺傳修飾生物體,如轉基因和“基因敲除”動物以及轉基因植物。
重組DNA技術已經對生物學和醫學產生巨大影響,並且由於整個人類基因組的核酸序列已經被了解,極可能會產生更大的影響。成千上萬種未知功能的基因將採用重組DNA技術來進行研究。基因治療可能成為某些疾病的常規療法,採用遺傳工程技術將可以設計出許多新的基因轉移載體。
同樣地,採用重組DNA技術獲得的轉基因植物將可能在現代農業中扮演日益重要的角色。涉及到構建或使用GMOs的實驗應首先進行生物安全評估。與該生物體有關的病原特性和所有潛在危害可能都是新型的,沒有確定的。供體生物的特性、將要轉移的DNA序列的性質、受體生物的特性以及環境特性等都需要進行評估。這些因素將有助於決定安全操作目標遺傳修飾生物體所要求的生物安全水平,並確定應使用的生物學和物理防護系統。
·生物表達系統的生物安全考慮
生物表達系統由載體和宿主細胞組成。必須滿足許多標準使其能有效、安全地使用。質粒pUC18是這樣一種生物表達系統的實例。質粒pUC18經常與大腸桿菌K12細胞一起使用作為克隆載體,其完整測序已經完成。所有需要在其他細菌表達的基因已經從它的前體質粒pBR322中刪除。大腸桿菌K12是一種非致病性菌株,它不能在健康人和動物的消化道中持久克隆。如果所要插入的外源DNA表達產物不要求更高級別的生物安全水平,那么大腸桿菌K12/pUC18可以在一級生物安全水平下按常規的遺傳工程實驗進行。
·表達載體的生物安全考慮
下列情況需要較高的生物安全水平:
1、來源於病原生物體的DNA序列的表達可能增加GMO的毒性
2、插入的DNA序列性質不確定,例如在製備病原微生物基因組DNA庫的過程中
3、基因產物具有潛在的藥理學活性
4、毒素的基因產物編碼。
·用於基因轉移的病毒載體
病毒載體(腺病毒載體)可以用於將基因有效地轉移到其他細胞。這樣的載體缺少病毒複製的某些基因,可以在能夠補充這些缺陷的細胞株內繁殖。這類病毒載體的貯存液中可能污染了可複製病毒,它們是由繁殖細胞株中極少發生的自發性重組產生的。這些載體操作時應採用與用於獲得這些載體的母體腺病毒相同的生物安全水平。
·轉基因動物和“基因敲除”動物
攜帶外源性遺傳信息的動物(轉基因動物)應當在適合外源性基因產物特性的防護水平下進行操作。特定基因被有目的地刪除的動物(“基因敲除”動物)一般不表現特殊的生物危害。包括那些表達病毒受體的轉基因動物一般不會感染該種系病毒。如果這種動物從實驗室逃離並將轉移基因傳給野生動物群體,那么理論上可以產生儲存這些病毒的動物宿主。
目前已經就脊髓灰質炎病毒,特別是與根除脊髓灰質炎相關的問題討論了上述可能性。由不同實驗室獲得的表達人脊髓灰質炎病毒受體的轉基因小鼠,它們對不同接種途徑的脊髓灰質炎病毒的感染都很敏感,所產生的疾病在臨床和組織病理學上也與人脊髓灰質炎相類似。但小鼠模型與人不同的是,在口腔接種脊髓灰質炎病毒後,腸道內的病毒複製不充分或沒有發生。因此,如果這種轉基因小鼠逃到野外,幾乎不可能產生脊髓灰質炎病毒新的宿主動物。但是,這個例子表明,對於每一種新的轉基因動物,應當通過詳細研究來確定動物的感染途徑、感染所需的病毒接種量以及感染動物傳播病毒的範圍。此外,應當採取一切措施以確保對受體轉基因小鼠的嚴密防護。
·轉基因植物
那些表達了能夠耐受除草劑或抵抗昆蟲能力等基因的轉基因植物,目前在世界許多地區都引起相當的爭議。這些爭議的焦點是這類植物作為食物的安全性,以及種植後的長期生態後果。表達動物或人源性基因的轉基因植物用於研發醫學產品和營養物品。通過危險度評估可以確定這些轉基因植物產品所需的生物安全水平。
