
信使介紹
Messenger RNA (mRNA)—— 信使核糖核酸
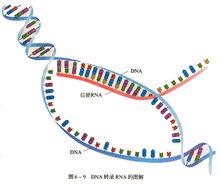 信使RNA
信使RNA攜帶遺傳信息,在蛋白質合成時充當模板的RNA。從脫氧核糖核酸(DNA)轉錄合成的帶有遺傳信息的一類單鏈核糖核酸(RNA)。它在核糖體上作為蛋白質合成的模板,決定肽鏈的胺基酸排列順序。mRNA存在於原核生物和真核生物的細胞質及真核細胞的某些細胞器(如線粒體和葉綠體)中。
原核生物和真核生物mRNA有不同的特點:
①原核生物mRNA常以多順反子的形式存在。真核生物mRNA一般以單順反子的形式存在。
②原核生物mRNA的轉錄與翻譯一般是偶聯的,真核生物轉錄的mRNA前體則需經轉錄後加工,加工為成熟的mRNA與蛋白質結合生成信息體後才開始工作。
③原核生物mRNA半壽期很短,一般為幾分鐘 ,最長只有數小時(RNA噬菌體中的RNA除外)。真核生物mRNA的半衰期較長, 如胚胎中的mRNA可達數日。
④原核與真核生物mRNA的結構特點也不同。
原核生物mRNA一般5′端有一段不翻譯區,稱前導區,3′端有一段不翻譯區,中間是蛋白質的編碼區,一般編碼幾種蛋白質。真核生物mRNA(細胞質中的)一般由5′端帽子結構、5′端不翻譯區、翻譯區(編碼區)、3′端不翻譯區和3′端聚腺苷酸尾巴構成。分子中除m7G構成帽子外,常含有其他修飾核苷酸,如m6A等。真核生物mRNA通常都有相應的前體。從DNA轉錄產生的原始轉錄產物可稱作原始前體(或mRNA前體)。一般認為原始前體要經過hnRNA核不均一RNA的階段,最終才被加工為成熟的mRNA。
通常mRNA(單鏈)分子自身回折產生許多雙鏈結構。原核生物,經計算有66.4%的核苷酸以雙鏈結構的形式存在。真核生物mRNA也具有豐富的二級結構,摺疊起來的mRNA二級結構有利於蛋白質合成的啟動,以後mRNA處於伸展的狀態則有利於轉譯的繼續。
mRNA的複製,轉錄和翻譯:由一個DNA分子,邊解旋,邊轉錄。利用細胞核內部的游離核糖核苷酸合成。合成規則遵循鹼基互補配對原則。註:因為mRNA沒有T(胸腺嘧啶),所以模版中出現A(腺嘌呤)時,由U(尿嘧啶)代替。以上過程叫做轉錄,在細胞核中完成。接著,mRNA穿過核孔。和細胞質中的核糖體結合。選擇tRNA運輸胺基酸,和對應的三個鹼基排列好(如何排列請查詢:密碼子)。再與其它的胺基酸通過肽鍵連線在一起,形成肽鏈。以上過程叫做翻譯,在細胞質中完成。
雖然人們已經破譯了決定生命基礎的蛋白質的胺基酸合成密碼,也知道了是DNA攜帶著這種密碼,但是,根據細胞學所掌握的事實:所有DNA都呆在細胞核內,而蛋白質卻存在於細胞質中,像DNA這樣的大分子是無法隨意進入細胞質的。然而密碼總是會被帶入細胞質的,這一來,人們不禁要問,是誰把鎖在細胞核內的DNA手裡的密碼帶入了細胞質的呢?科學家們從DNA那裡拷貝了一份密碼檔案,並帶入了細胞質中。經過試驗和觀察,發現這個信使就是RNA——核糖核酸。
發現
儲存在DNA分子中的這種遺傳信息能在複製中產生更多的拷貝,並翻譯成蛋白質。DNA的功能構成了信息的流動,遺傳信息如何轉變成蛋白質呢?轉錄就是其中的重要的一環。基因表達時以DNA的一條鏈為模板合成RNA,這一過程就是轉錄(transcription)。催化合成RNA的酶叫做RNA聚合酶(RNA polymerase)。RNA和DNA結構相似,所不同之處在於:⑴RNA一般以單鏈形式存在;⑵RNA中的核糖其C′-2不脫氧;⑶尿苷(U)取代了DNA中的胸苷。細胞中的RNA分成三種:mRNA(信使RNA),tRNA(轉運RNA)和rRNA(核糖體RNA)。它們的功能各不相同。mRNA是合成蛋白質的模板,tRNA是轉運特異胺基酸的運載工具,rRNA是合成蛋白質的裝置。mRNA的鹼基序列,決定著蛋白質裝配時胺基酸的序列。
1955年Brachet用洋蔥根尖和變形蟲進行了實驗;若加入RNA酶降解細胞中的RNA,則蛋白質合成就停止,若再加入從酵母中提取的RNA,則又可以重新合成一些蛋白質,這就表明,蛋白質的合成是依賴於RNA。
同年Goldstein和Plaut用同位素標記變形蟲(Amoeba proteus)RNA前體,發現標記的RNA都在核內,表明RNA是在核內合成的。在標記追蹤(pulse-chase)實驗中,用短脈衝標記RNA前體,然後將細胞核轉移到未標記的變形蟲中。經過一段時間發現被標記的RNA分子已在細胞質中,這就表明RNA在核中合成,然後轉移到細胞質內,而蛋白質就在細胞質中合成,因此RNA就成為在DNA和蛋白質之間傳遞信息的信使的最佳候選者。
1956年Elliot Volkin和 Lawrence Astrachan作了一項很有意思的觀察:當E.coli被T2感染,迅速停止了RNA的合成,但噬菌的RNA卻開始迅速合成。用同位素脈衝一追蹤標記表明噬菌的RNA在很短的時間內就進行合成,但很快又消失了,表明RNA的半衰期是很短的。由於這種新合成的RNA的鹼基比和T2的DNA鹼基比相似,而和細菌的鹼基比不同,所以可以確定新合成的RNA是T2的RNA。由於T2感染細菌時注入的是DNA,而在細胞里合成的是RNA,可見DNA是合成RNA的模板。最令人信服的證據來自DNA-RNA的雜交實驗。Hall.B.D和Spiegeman,S,將T2噬菌體感染E.coli後立即產生的RNA分離出來,分別與T2和E.coli的DNA進行分子雜交,結果發現這種RNA只能和T2的DNA雜交形成“雜種”鏈,而不能和E.coli的DNA進行雜交。表明T2產生的這種RNA(即mRNA)至少和T2的DNA中的一條鏈是互補的。
Brenner,s. Jacob,F.和Meselson(1961)進行了一系列的的實驗(圖12-2),他們將E.coli培養在15N/13C的培養基中,因此合成的RNA和蛋白都被“重”同位素所標記。也就是說凡是“重”的核糖體,RNA和蛋白都是細菌的,然後用T2感染E.coli,細菌的RNA停止合成,而開始合成T2的RNA此時用普通的“輕”培養基(14N/12C),但分別以32P來標記新合成的T2 RNA,以35S標記新合成的T2蛋白,因此任何重新合成的核糖體,RNA,及蛋白都是“輕”的但帶但有放射性同位素。經培養一段時間後破碎細胞,加入過量的輕的核糖體作對照,進行密度梯度離心,結果“輕”的核糖體上不具有放射性,“重”的核糖體上具有32P和35S,表明⑴T2未合成核糖體,“輕”核糖體卻是後加放的。⑵T2翻譯時是借用了細菌原來合成的核糖體,所以核糖體並無特異性,核糖體上結合的mRNA,其序列的特異性才是指導合成蛋白質的遺傳信息,從而提出了mRNA作為“信使”的證據。因此他們將這種能把遺傳信息從DNA傳遞到蛋白質上的物質稱為“信使”。他們預言⑴這種“信使”應是一個多核苷酸;⑵②其平均分子量不小於5´105(假定密碼比是3),足以攜帶一個基因的遺傳信息;⑶它們至少是暫時連在核糖體上;⑷其鹼基組成反映了DNA的序列;⑸它們能高速更新。Volkin和Astrachan發現高速更新的RNA似乎完全符合以上條件。Jacob和Monod將它定名為信使RNA(Messenger RNA)或mRNA。
合成與加工
第一節 DNA轉錄生成RNA
一、定義
一轉錄單位
二啟動子(promoter)
三終止子(terminator)
二、RNA聚合酶
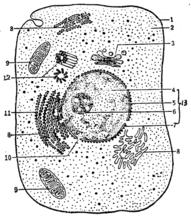 以DNA為模板形成信使RNA的過程
以DNA為模板形成信使RNA的過程一酶的特性:以4種NTP為底物,需模板和鎂離子,合成方向也是5’-3’,但不需要引物。
二酶的分類:
1.噬菌體的RNA聚合酶結構簡單,是單鏈蛋白,功能也簡單。
2.細菌則具有複雜的多亞基結構(450Kd),可識別並轉錄超過1000個轉錄單位。
3.真核生物的酶有多種,根據a-鵝膏蕈鹼(環狀8肽,阻斷RNA延伸)的抑制作用可分為三類:聚合酶A對它不敏感,分布於核仁,轉錄核糖體RNA;聚合酶B對低濃度敏感,存在於核質,轉錄信使RNA;聚合酶C位於核質,對高濃度敏感,轉錄小分子量RNA,如轉運RNA、5SRNA等。各種RNA聚合酶都是由10-15種不同亞基組成的多亞基複合物。
4. 線粒體和葉綠體也有RNA聚合酶,結構簡單,能合成所有種類RNA。
三酶的構成:大腸桿菌的全酶有5個亞基(α2ββ’ωσ),含2個鋅。β催化形成磷酸二酯鍵,β’結合模板,σ亞基稱為起始因子,可使RNA聚合酶穩定地結合到啟動子上。ββ’ωσ稱為核心酶。σ亞基在不同菌種間變動較大,而核心酶比較恆定。酶與不同啟動子的結合能力不同,不同啟動因子可識別不同的啟動子。σ70識別啟動子共有序列,σ32識別熱休克基因,σ60在氮飢餓時起作用。σ通過隨機移動起作用,不需解鏈。
四模板:以完整雙鏈DNA為模板,其中僅一條鏈可轉錄。轉錄時局部解鏈,轉錄後DNA重新形成雙螺旋結構,所以DNA是全保留的。
三、轉錄過程
分為起始、延長和終止三個階段。起始包括對雙鏈DNA特定部位的識別、局部(17bp)解鏈以及在最初兩個核苷酸間形成磷酸二酯鍵。第一個核苷酸摻入的位置稱為轉錄起點。
起始後起始因子離開,核心酶構象改變,沿模板移動,轉錄生成雜交雙鏈(12bp)。隨後DNA互補鏈取代RNA鏈,恢復DNA雙螺旋結構。延伸速度為50nt/s,酶移動17nm。錯誤幾率為10-5。
聚合酶到達終點時,在終止輔助因子的幫助下停止反應,酶和RNA鏈脫落,轉錄結束。
四、啟動子和轉錄因子
一定義:酶識別、結合、開始轉錄的一段DNA序列。強啟動子2秒鐘啟動一次轉錄,弱啟動子10分鐘一次。
二原核生物:大腸桿菌在起點上游約-10鹼基對處有保守序列TATAAT,稱為pribnow box,有助於局部解鏈。在其上游還有TTGACA,稱為-35序列,提供RNA聚合酶識別的信號。
三真核生物:複雜,差異較大。
1.信使RNA的啟動子通常有三個保守區,-25到-30有TATA框,是解鏈位置,並決定轉錄起點;-75位置有CAAT框,與RNA聚合酶的結合有關;更上游還有GC框,某些轉錄因子可結合。後兩個稱為上游因子,對轉錄起始頻率有較大影響;
2. 小RNA的啟動子在轉錄區內部,有一些輔助因子幫助RNA聚合酶識別。
五、終止子和終止因子
一定義
二所有原核生物的終止子在終點之前都有一個迴文結構,可使酶減慢移動或暫停合成。大腸桿菌有兩類終止子:
1. 簡單終止子,回文區有一段富含GC對的序列,回文後有寡聚尿苷。
2.依賴ρ的終止子,必須在有ρ因子時才能發揮作用,不含GC對,也無寡聚尿苷。ρ因子是蛋白質,可與酶作用,釋放RNA,並使酶脫離。
三某些因子可使酶越過終止子繼續轉錄,稱為通讀。常見於某些噬菌體的時序控制,早期基因與晚期基因以終止子相隔,早期基因產生抗終止因子,使發生通讀以表達晚期基因。
六、轉錄的調控
一遺傳信息的表達有時序調控和適應調控,轉錄水平的調控是關鍵環節,因為這是表達的第一步。轉錄調控主要發生在起始和終止階段。
二操縱子是細菌基因表達和調控的單位,有正調節和負調節因子。阻遏蛋白的作用屬於負調控。環腺苷酸通過其受體蛋白(CRP)促進轉錄,可促進許多誘導酶的合成。操縱子可構成綜合性調控網路,如SOS反應等。對終止子也有調控作用,如衰減子。
三真核生物不組成操縱子,而是通過激素、生長因子等進行調控。某些DNA序列對轉錄起增強作用,稱為增強子。
第二節 轉錄後加工
一、原核生物
一核糖體RNA:大腸桿菌共有7個核糖體RNA的轉錄單位,每個轉錄單位由16S、23S、5SRNA和若干轉運RNA基因組成。16S和23S之間常由轉運RNA隔開。轉錄產物在RNA酶III的作用下裂解產生核糖體RNA的前體P16和P23,再由相應成熟酶加工切除附加序列。前體加工時還進行甲基化,產生修飾成分,特別是a-甲基核苷。N4,2’-O二甲基胞苷(m4Cm)是16S核糖體RNA特有成分。5S核糖體RNA一般無修飾成分。
二轉運RNA:有60個基因,其加工包括:
1.內切酶在兩端切斷,大腸桿菌RNA酶P是5’成熟酶;
2.外切酶從3’修剪,除去附加順序。RNA酶D是3’成熟酶;
3.3’端加上CCAOH,由轉運RNA核苷醯轉移酶催化,某些轉運RNA已有,切除附加序列後即露出;
4.核苷的修飾:修飾成分包括甲基化鹼基和假尿苷,修飾酶具有高度特異性。甲基化對鹼基和序列都有嚴格要求,一般以S-腺苷甲硫氨酸為甲基供體。
三信使RNA:細菌多數不用加工,轉錄與翻譯是偶聯的。也有少數多順反子信使RNA必須由內切酶切成較小的單位,然後翻譯。如核糖體大亞基蛋白與RNA聚合酶的b亞基基因組成混合操縱子,轉錄後需經RNA酶III切開,各自翻譯。因為RNA聚合酶的合成水平低得多,切開有利於各自的翻譯調控。較長的RNA會產生高級結構,不利於翻譯,切開可改變其結構,從而影響其功能。
二、真核生物
一核糖體RNA:基因拷貝數多,在幾十到幾千之間。基因成簇排列在一起,由RNA聚合酶I轉錄生成一個較長的前體,哺乳動物為45S。核仁是rRNA合成與核糖體亞基生物合成的場所。 RNA酶III等核酸內切酶在加工中起重要作用。5SRNA基因也是成簇排列的,由RNA聚合酶III轉錄,經加工參與構成大亞基。核糖體RNA可被甲基化,主要在核苷2’羥基,比原核生物甲基化程度高。多數核糖體RNA沒有內含子,有些有內含子但不轉錄。
二轉運RNA:由RNA聚合酶III轉錄,加工與原核相似,但3’端的CCA都是後加的,還有2’-O-甲基核糖。
三信使RNA:真核生物編碼蛋白質的基因以單個基因為轉錄單位,但有內含子,需切除。信使RNA的原初轉錄產物是分子量很大的前體,在核內加工時形成大小不等的中間物,稱為核內不均一RNA(hnRNA)。其加工過程包括:
1.5’端加帽子:在轉錄的早期或轉錄終止前已經形成。首先從5’端脫去一個磷酸,再與GTP生成5’,5’三磷酸相連的鍵,最後以S-腺苷甲硫氨酸進行甲基化,形成帽子結構。帽子結構有多種,起識別和穩定作用。
2. 3’端加尾:在核內完成。先由RNA酶III在3’端切斷,再由多聚腺苷酸聚合酶加尾。尾與通過核膜有關,還可防止核酸外切酶降解。
3. 內部甲基化:主要是6-甲基腺嘌呤,在hnRNA中已經存在。可能對前體的加工起識別作用。
三、RNA的拼接
一轉運RNA的拼接:由酶催化,酶識別共同的二級結構,而不是序列。通常內含子插入到靠近反密碼子處,與反密碼子配對,取代反密碼子環。第一步由內切酶切除插入序列,不需ATP;第二步由RNA連線酶連線,需要ATP。
二四膜蟲核糖體RNA的拼接:某些四膜蟲26S核糖體RNA基因中有一個內含子,其拼接只需一價和二價陽離子及鳥苷酸或鳥苷存在即可自發進行。其實質是磷酸酯的轉移反應,鳥苷酸起輔助因子的作用,提供游離3’羥基。
三信使RNA:真核生物編碼蛋白質的核基因的內含子屬於第二類內含子,左端為GT,右端為AG。先在左端切開,產生的5’末端與3’端上游形成5’,2’-磷酸二酯鍵,構成套索結構。然後內含子右端切開,兩個外顯子連線起來。通過不同的拼接方式,可形成不同的信使RNA。
第三節 RNA的複製
一、噬菌體QbRNA的複製
其RNA是單鏈,正鏈,侵入大腸桿菌後立即翻譯,產生複製酶的b亞基,與宿主的三個亞基(α為核糖體蛋白,γ、δ均為肽鏈延長因子)構成複製酶,進行複製。先以正鏈為模板合成負鏈,再根據負鏈合成正鏈。合成負鏈時需要宿主的兩個蛋白因子,合成正鏈則不需要,所以可大量合成。病毒的蛋白質合成受RNA高級結構的調控。
二、病毒RNA複製的主要方式
一病毒含正鏈RNA,先合成複製酶,複製後合成其他蛋白質進行裝配。如噬菌體Qb及灰質炎病毒。
二病毒含負鏈和複製酶,先合成正鏈,再合成病毒蛋白和複製病毒RNA。如狂犬病毒。
三病毒含雙鏈RNA和複製酶,如呼腸孤病毒。先複製正鏈,再翻譯成病毒蛋白,最後合成負鏈,形成雙鏈RNA分子。
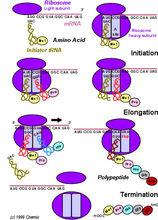 核糖體:多肽合成場所,能與信使RNA結合
核糖體:多肽合成場所,能與信使RNA結合四致癌RNA病毒:如白血病病毒和肉瘤病毒,先逆轉錄生成DNA前病毒,再轉錄、翻譯。
第四節 RNA生物合成的抑制劑
一、鹼基類似物
有些人工合成的鹼基類似物能幹擾和抑制核酸的合成。作用方式有以下兩類:
一作為代謝拮抗物,直接抑制核苷酸生物合成有關酶類。如6-巰基嘌呤進入體內後可轉變為巰基嘌呤核苷酸,抑制嘌呤核苷酸的合成。可作為抗癌藥物,治療急性白血病等。此類物質一般需轉變為相應的核苷酸才能表現出抑制作用。
二進入核酸分子,形成異常RNA或DNA,影響核酸的功能並導致突變。5-氟尿嘧啶類似尿嘧啶,可進入RNA,與腺嘌呤配對或異構成烯醇式與鳥嘌呤配對,使A-T對轉變為G-C對。因為正常細胞可將其分解,而癌細胞不能,所以可選擇性抑制癌細胞生長。
二、DNA模板功能抑制物
一烷化劑:帶有活性烷基,能使DNA烷基化。鳥嘌呤烷化後易脫落,雙功能烷化劑可造成雙鏈交聯,磷酸基烷化可導致DNA鏈斷裂。通常有較大毒性,引起突變或致癌。
二放線菌素類:可與DNA形成非共價複合物,抑制其模板功能。包括一些抗癌抗生素。
三嵌入染料:含有扁平芳香族發色團,可插入雙鏈DNA相鄰鹼基對之間。常含丫啶或菲啶環,與鹼基大小類似,可在複製時增加一個核苷酸,導致移碼突變。如溴乙啶。
三、RNA聚合酶抑制劑
一利福黴素:抑制細菌RNA聚合酶活性。
二利鏈菌素:與細菌RNA聚合酶b亞基結合,抑制RNA鏈的延長。
a-鵝膏蕈鹼:抑制真核生物RNA聚合酶。
結構與功能
從 (DNA)轉錄合成的帶有遺傳信息的一類單鏈(RNA),他作為蛋白質合成的模板,決定了核糖體合成肽鏈的種類。1961年F.雅各布和根據大腸桿菌誘導酶生成的實驗結果提出:信息從DNA到蛋白質之間的轉移,必需有一種RNA起傳遞作用,由此提出了信使核糖核酸的名稱。
生物體內的每種多肽鏈都由特定的mRNA編碼,所以細胞內mRNA的種類很多,但通常每種mRNA的拷貝數極少(1~10個)。根據信息密碼學說,3個連續的核苷酸可以編碼一個胺基酸,因此從已知mRNA(或DNA)核苷酸順序可以準確推導出蛋白質的一級結構。
存在範圍和性質
mRNA存在於原核和真核生物的細胞質及真核細胞的某些細胞器(如和)中。RNA病毒和RNA噬菌體中的 RNA既是遺傳信息的載體又具有mRNA的功能。生物體mRNA種類的多少與生物進化水平有關,高等生物所含的遺傳信息多,mRNA的種類也多。生物體內某種mRNA的含量根據需要而有不同,如5齡蠶後部絲腺體的主要任務是快速合成大量絲心蛋白,因而編碼絲心蛋白的mRNA含量特別多。有些細菌需要不斷適應外部環境,其體內編碼某些誘導酶的mRNA的含量也較多。
原核和真核生物mRNA不同的特點
①原核生物mRNA常以多順反子(見)的形式存在,即一條mRNA鏈編碼幾種功能相關聯的蛋白質。真核生物mRNA一般以單順反子的形式存在,即一種mRNA只編碼一種蛋白質。②原核生物mRNA的轉錄與翻譯一般是偶聯的,即轉錄尚未完畢,蛋白質的轉譯合成就已開始 真核生物轉錄的mRNA前體則需經後加工,加工為成熟的mRNA與蛋白質結合生成信息體後才開始工作 信息體中蛋白質與RNA之比約為3。③原核生物mRNA半壽期很短,一般為幾分鐘,最長只有數小時(RNA噬菌體中的RNA除外)。真核生物mRNA的半壽期較長,如胚胎中的mRNA可達數日。④原核與真核生物mRNA的結構特點也不同。
一級結構與功能的關係
原核生物mRNA一般5'端有一段不翻譯區,稱前導順序,3'端有一段不翻譯區,中間是蛋白質的編碼區,一般編碼幾種蛋白質。如大腸桿菌乳糖操縱子mRNA編碼3條多肽鏈;色氨酸操縱子mRNA編碼5條多肽鏈。也有單順反子形式的細菌mRNA,如大腸桿菌脂蛋白mRNA。原核生物mRNA分子中一般沒有修飾核苷酸,也沒有5'端帽子結構和3'端聚腺苷酸尾巴。在原核生物mRNA的起始密碼子(AUG)附近(5'方向上游)的一小段長短不等的順序,含有較多的嘌呤核苷酸,被稱為SD順序。它能和核糖體小亞基上的16SrRNA的3'端富含嘧啶核苷酸的區域配對結合,有助於帶有甲醯甲硫氨酸的起始tRNA識別mRNA上的起始密碼(AUG),使肽鏈合成從此開始。這段順序是1974年由J.夏因和L.達爾加諾發現的,所以稱為SD順序,也稱核糖體結合部位。原核生物mRNA的編碼區一般編碼幾種功能上相關聯的蛋白質,兩種蛋白質的編碼區之間常有一小段不翻譯的順序,叫做間隔區。有的噬菌體RNA中2個相鄰的順反子共用一段相同的編碼順序,例如,M 噬菌體RNA中的溶菌蛋白編碼區共225個核苷酸中有189個核苷酸是由相鄰兩個蛋白質共用的。原核mRNA與真核mRNA一樣使用同一套三聯體密碼子(真核生物線粒體mRNA有例外)。原核生物合成胺基酸的操縱子mRNA的5' 端前導順序上有一段順序稱作弱化子。弱化子具有兩種可以互變的構象,其中一種構象是轉錄終止的信號,能使轉錄中止(或衰減)。衰減調節是原核生物合成胺基酸的調控方式之一(見)。
真核生物 mRNA(細胞質中的)一般由5'端帽子結構、5'端不翻譯區、翻譯區(編碼區)、3'端不翻譯區和3'端聚腺苷酸尾巴構成(圖1a[真核生物mRNA結構示意圖 a一級結構示意圖])。分子中除 G構成帽子外,常含有其他修飾核苷酸,如 A等。5'端帽子結構通常有3種類型,即:G(5')ppp(5')N; G(5')ppp(5') N和 G(5')ppp(5') N。圖1b[真核生物mRNA結構示意圖b 5'端帽子結構式,,表示鹼基],表示鹼基" class=image>[] 是帽子的化學結構,N右邊的m代表核糖2'位羥基的甲基化。真核細胞線粒體中的mRNA無帽子結構。一般認為帽子的功能與翻譯的啟動有關。許多真核生物 mRNA(如珠蛋白mRNA)除去帽子後翻譯效率大大降低。5'端不翻譯區,也叫前導順序。不同的真核mRNA的前導順序長度不同,有的只有10個核苷酸,有的則有200個核苷酸。與原核mRNA相似,真核mRNA5'端不翻譯區中常有一段順序與核糖體小亞基上的18SrRNA的3'端的一段順序互補並結合,這種結合與真核mRNA的翻譯啟動有關。
翻譯區(編碼區)使用的密碼子除線粒體(如人、牛和酵母線粒體)外與原核生物mRNA是一樣的。真核生物mRNA的起始密碼子都是AUG。真核和原核生物mRNA使用的密碼子也都有“簡併現象”,即幾種不同的密碼子翻譯出同一種胺基酸,但不同的mRNA中簡併密碼子的利用率是不同的,真核與原核生物之間的差別就更大。mRNA的終止密碼子有3個(UAG、UGA和UAA),其功能是停止翻譯,一般只用一個終止密碼子就能使翻譯停止。有的mRNA有2個連續的終止密碼子(見)。3'端不翻譯區的長短在不同的mRNA上有所不同,β珠蛋白mRNA只有39個核苷酸,而卵白蛋白mRNA則有637個核苷酸。真核生物mRNA3'端不翻譯區常有 AAUAA(A)或AUUUA(A)等順序,它們和識別多聚A聚合酶及裝配多聚A尾巴有關。除個別組蛋白mRNA外,真核生物mRNA3'端均有多聚A尾巴 3'端多聚A尾巴的長度隨來源不同而不同,且隨mRNA的老化而變短,通常有20~200個A。多聚A與mRNA穩定性及mRNA從細胞核轉到細胞漿中有關。
真核生物mRNA的前體 真核生物mRNA通常都有相應的前體。從DNA轉錄產生的原始轉錄產物可稱作原始前體(或mRNA前體)。一般認為原始前體要經過hnRNA核不均-RNA的階段,最終才被加工為成熟的 mRNA。hnRNA上的蛋白質編碼區被一些居間順序分隔成若干段;不同的基因轉錄產物所含的居間順序的數目不同,人胰島素只有兩個,而牛眼的晶體蛋白則含有數十個;居間順序的長短也各不相同,從數十個到上千個核苷酸(雞卵白蛋白有一個1550個核苷酸的居間順序)。居間順序將在剪接過程中去除。約有10~40%的hnRNA含有3′端多聚A尾巴。hnRNA經過進一步加工切除居間順序並把分隔的蛋白質編碼區連線起來,最終成為成熟的mRNA。
二級結構與功能的關係
 DNA表達的信息需轉給一種“信使RNA”
DNA表達的信息需轉給一種“信使RNA”通常mRNA(單鏈)分子自身回折產生許多雙鏈結構(圖2 [噬菌體M RNA中成熟蛋白] RNA中成熟蛋白" class=image>[編碼區的二級結構及外殼蛋白的起始密碼子 AUG的位置])。原核生物,例如M 噬菌體RNA外殼蛋白編碼區,經計算有66.4%的核苷酸以雙鏈結構的形式存在。M RNA能翻譯4種蛋白質,但效率各不相同。在通常條件下翻譯外殼蛋白(其編碼區在成熟蛋白的下游)的效率高於成熟蛋白的效率。但用甲醛處理M RNA破壞二級結構後,則翻譯成熟蛋白的效率提高。圖2[噬菌體M RNA中成熟蛋] RNA中成熟蛋" class=image>[白編碼區的二級結構及外殼蛋白的起始密碼子 AUG的位置] 中外殼蛋白的起始密碼子 AUG(1335~1337)通常處於環(Loop)的頂端,暴露在外面,因而易於與翻譯的啟動因子結合而進行翻譯。成熟蛋白的編碼區儘管處在外殼蛋白的前面,但其起始密碼子GUG(130~132)卻埋在二級結構之中,故翻譯效率低,只有將二級結構鬆開(如甲醛處理)之後才能被翻譯。可見mRNA分子的二級結構對翻譯蛋白質的效率有很大影響。
真核生物mRNA也具有豐富的二級結構,如鴨珠蛋白mRNA和兔珠蛋白mRNA分別有45~60%和55~62%的核苷酸殘基處在鹼基配對之中。在真核生物蛋白質啟動複合物中,40S核糖體實際上覆蓋著mRNA上包括帽子結構在內的50~54個核苷酸,但是40S核糖體的大小比50個核苷酸的長度小得多 由於形成的髮夾結構(二級結構使帽子與起始密碼子之間的空間距離縮短)(圖3[真核生物mRNA ]),造成40S核糖體能夠覆蓋包括帽子結構和起始密碼子 AUG在內的50多個核苷酸,從而啟動蛋白質合成。不同的mRNA中髮夾結構的有無或多少各不相同。在蛋白質合成肽鏈繼續延伸時,不需要帽子結構參加,此時核糖體覆蓋的mRNA的區域約為25~35個核苷酸,mRNA的構象已不同於啟動階段而是處於一種伸展的狀態,從而有利於轉譯的延續。可見,摺疊起來的mRNA二級結構有利於蛋白質合成的啟動,以後mRNA處於伸展的狀態則有利於轉譯的繼續。
信使與密碼子
簡介
遺傳信息從DNA分子抄錄到RNA分子中的過程稱為轉錄(transcription)。在真核生物中,最初轉錄生成的RNA稱為核不均一RNA(heterogeneous nuclear RNA,hnRNA),然而在細胞漿中起作用,作為蛋白質的胺基酸序列合成模板的是mRNA(messenger RNA)。hnRNA是mRNA的未成熟前體。兩者之間的差別主要有兩點:一是hnRNA核苷酸鏈中的一些片段將不出現於相應的mRNA中,這些片段稱為內含子(intron),而那些保留於mRNA中的片段稱為外顯子(exon)。也就是說,hnRNA在轉變為mRNA的過程中經過剪接,被去掉了一些片段,餘下的片段被重新連線在一起;二是mRNA的5′末端被加上一個m7pGppp帽子,在mRNA3′末端多了一個多聚腺苷酸(polyA)尾巴。mRNA從5′末端到3′末端的結構依次是5′帽子結構,5′末端非編碼區,決定多肽胺基酸序列的編碼區,3′末端非編碼區,和多聚腺苷酸尾巴。多聚腺苷酸尾一般由數十個至一百幾十個腺苷酸連線而成。隨著mRNA存在時間的延續,這段聚A尾巴慢慢變短。因此,目前認為這種3′末端結構可能與增加轉錄活性以及使mRNA趨於相對穩定有關。原核生物的mRNA沒有這種首、尾結構。
1961年,Jacob和Monod首先提出了mRNA的概念。在真核細胞中,由於蛋白質是在胞漿中而不是在核內合成,因此顯然要求有一個中間物將DNA上的遺傳信息傳遞至胞漿中。後來的研究證實,這種中間物即信使RNA.?mRNA的核苷酸序列與DNA序列相應,決定著合成蛋白質的胺基酸序列。它如何指導胺基酸以正確的順序連線起來呢?不同的mRNA鹼基組成和排列順序都不同,但都只有A,G,C,U4種鹼基。如果一個鹼基就可以決定一個胺基酸,則只有四種變化方式,如果兩個鹼基決定一個胺基酸,則只有16種變化方式,都不能滿足20種胺基酸的需要。1961年Crick和Brenner的實驗得出了三個核苷酸編碼一個胺基酸的結論,並將這種三位一體的核苷酸編碼稱做遺傳密碼(genetic code)或三聯體密碼,這樣就可以有64種不同的密碼,但此情況下必須假定有一些胺基酸使用兩個以上的密碼。這一假定很快就被證明是對的。
遺傳密碼具有下列特徵:
1.三個核苷酸組成一個密碼子,每個密碼子由三個前後相聯的核苷酸組成,一個密碼子只為一種胺基酸編碼。共有64個密碼子;
2.密碼子之間不重疊使用核苷酸,也無核苷酸間隔;
3.一種胺基酸可有多個密碼子,這個特點稱為密碼子的簡併性;
4.密碼子的通用性,所有生物從最低等的病毒直至人類,蛋白質合成都使用同一套密碼子表(表15-8),僅有極少的例外,如特殊細胞器線粒體,葉綠體所用的密碼稍有不同。
通用遺傳密碼及相應的胺基酸
第一個核苷酸5′ 第二個核苷酸 第三個核苷酸3′
U C A G
U 苯丙氨酸 絲氨酸 酪氨酸 半胱氨酸 U
苯丙氨酸 絲氨酸 酪氨酸 半胱氨酸 C
亮氨酸 絲氨酸 終止碼 終止碼 A
亮氨酸 絲氨酸 終止碼 色氨酸 G
C 亮氨酸 脯氨酸 組氨酸 精氨酸 U
亮氨酸 脯氨酸 組氨酸 精氨酸 C
亮氨酸 脯氨酸 谷氨醯胺 精氨酸 A
亮氨酸 脯氨酸 谷氨醯胺 精氨酸 G
A 異亮氨酸 蘇氨酸 天冬醯胺 絲氨酸 U
異亮氨酸 蘇氨酸 天冬醯胺 絲氨酸 C
異亮氨酸 蘇氨酸 賴氨酸 精氨酸 A
蛋氨酸 蘇氨酸 賴氨酸 精氨酸 G
G 纈氨酸 丙氨酸 天冬氨酸 甘氨酸 U
纈氨酸 丙氨酸 天冬氨酸 甘氨酸 C
纈氨酸 丙氨酸 谷氨酸 甘氨酸 A
纈氨酸 丙氨酸 谷氨酸 甘氨酸 G
通用遺傳密碼與線粒體遺傳密碼之間的一些差異
密碼子 通用編碼 線粒體編碼
哺乳動物 果蠅 酵母菌 植物
UGA 終止碼 色氨酸 色氨酸 色氨酸 終止碼
AUA 異亮氨酸 蛋氨酸 蛋氨酸 蛋氨酸 異亮氨酸
CUA 亮氨酸 亮氨酸 亮氨酸 蘇氨酸 亮氨酸
AGA 精氨酸 終止碼 絲氨酸 精氨酸 精氨酸
註:下標橫線者為與通用編碼不同的編碼
究竟哪一個密碼子為哪一種胺基酸編碼,即密碼子與胺基酸之間的對應關係已在60年代研究解決了。1964年Nirenberg用一種RNA聚合酶體外合成了多聚尿苷酸、多聚腺苷酸等多聚核苷酸,將這些多聚核苷酸分別用於蛋白質的體外合成。發現,當所用的多聚核苷酸為多聚尿苷酸時,只有多聚苯丙氨酸合成,這意味著UUU為苯丙氨酸編碼;用其它多聚核苷酸進行相應的實驗後發現,CCC為脯氨酸編碼,而AAA為賴氨酸編碼;其後,有人又用核苷酸比例為已知,但是核苷酸序列隨機的多聚核苷酸,以及用已知序列的含兩種或兩種以上核苷酸的多聚核苷酸進行相應的實驗,將結果加以數理統計處理,又解讀了一批密碼子,其中包括三個終止碼,最後,還有一些密碼子是通過合成已知序列的三聚核苷酸與核蛋白體和載有放射性同位素標記的胺基酸的tRNA共沉澱原理予以解讀的。在所有密碼子中,AUG不僅為蛋氨酸編碼,而且又是翻譯(translation,以mRNA上的遺傳信息指導核蛋白體上多肽鏈合成的過程)的起始信號,UAA、UAG和UGA不為任何胺基酸編碼,而是作為翻譯的終止信號,統稱為終止碼(stop codon),又常被叫作無意義碼(nonsense codon)。
大多數胺基酸是由一個以上的密碼子所編碼。這個事實提出了一個問題:編碼同一種胺基酸的一組密碼子的使用頻率是否都相同?細緻的分析表明,無論是原核生物,還是高等真核生物,密碼子的使用頻率並不是平均的,有些密碼子的使用率很高,有些則幾乎不使用,其使用頻率主要與細胞內tRNA含量呈正相關。
傳遞
簡介
轉錄是在原核和真核細胞中以DNA為模板合成RNA的過程。
在原核和真核生物中,轉錄過程是相似的。包括DNA變性,RNA聚合酶結合在單鏈DNA上以5′→3′方向合成RNA分子。雙鏈中只有一條鏈作為轉錄模板,合成單鏈RNA分子。啟動子和終止子序列決定轉錄的起始和終止。
在E.coli中RNA多聚酶轉錄各種RNA(mRNA,tRNA和rRNA)。在真核細胞中有三類不同的RNA多聚酶,它們的功能不同。RNA pol Ⅰ轉錄4種rRNA中的3種;RNA pol Ⅱ轉錄mRNA和一些snRNA;RNAⅢ轉錄第四種rRNA,tRNA以及其餘的snRNA。
3種真核生物的RNA pol,不像E.coli RNA pol,沒有一個直接地和啟動子區結合,而是通過轉錄起始因子的介導來起始RNA的合成。對於每一種RNA多聚酶來說,轉錄因子是特異的,它可以識別啟動子的特殊序列。
蛋白質編碼基因的啟動子位於轉錄起始位點的上游,由不同組合的啟動原件所構成。特異的轉錄因子和調節因子結合在這些原件上,促進RNA pol Ⅱ轉錄起始。增強子離啟動子較遠,它可被調節因子識別結合,具有促進基因轉錄的功能。
由RNA pol Ⅲ轉錄的啟動子,位於下游,在其基因編碼序列內部。這種啟動子,根據所轉錄的RNA的種類,由不同的功能區組合而構成。轉錄因子識別這些功能區,促進RNA聚合酶轉錄起始。
18S,5.8S和28S rRNA作為一個轉錄單位一道轉錄,產生前體RNA分子。大部分真核生物的18S,5-8S和28S rRNA都是以串聯重複排列,每個重複單位被不轉錄的間隔序列(nontranscribed specer,NTs)所分隔。轉錄單位的啟動子位於NTS中,其功能是和特異的轉錄因子相結合,促進RNA pol Ⅰ的轉錄起始。
中心法則的提出
從孟德爾定律問世以後,人們就知道了生物的各種性狀是由基因控制的。一基因一酶學說的建立進一步地明確了基因是以酶的形式通過控制生化反應鏈來控制的。酶或蛋白和基因又是什麼樣的關係呢?也就是說遺傳信息怎樣傳遞,怎樣表達成性狀呢?就在Watson和Crick建立DNA雙螺旋模型後的第三年,1957年Crick提出了中心法測(central dogma),指出了遺傳信息的傳遞方向:
DNA → RNA→蛋白質
1970年H.Temin和D.Baltimore發現了反轉錄酶後,Crick對中心法測又作了部分修改:
DNA RNA →蛋白質
也就是說由DNA通過轉錄將遺傳信息傳遞給RNA,RNA通過翻譯把信息傳遞給蛋白。通過這種單向的傳遞,遺傳信息通過蛋白質的不同形式,如酶,結構蛋白,運載蛋白,調節蛋白等表達成一種性狀。
與轉運區別
區分mRNA和tRNA,可以從結構和功能這兩個方面去把握。
結構
mRNA的一級結構,tRNA的二級、三級結構是經常考察的內容,需要仔細區分。
⑴真核生物的mRNA的5' 端有帽子結構,3' 端為多聚腺苷酸(poly(A))尾巴。
⑵tRNA的二級結構呈三葉草形。三葉草形結構由胺基酸臂、二氫尿嘧啶環、反密碼環、額外環和TφC環等5個部分組成。其中,胺基酸臂末端為CCA;反密碼環中部為反密碼子,由3個鹼基組成。反密碼子可識別mRNA的密碼子。
⑶tRNA摺疊形成三級結構。tRNA的三級結構呈倒L形,反密碼環和胺基酸臂分別位於倒L的兩端。
功能
⑴mRNA是合成蛋白質的直接模板。每一種多肽鏈都有一種特定的mRNA做模板,因此細胞內mRNA的種類也是很多的。它將DNA上的遺傳信息轉錄下來,攜帶到核糖體上,在那裡以密碼的方式控制蛋白質分子中胺基酸的排列順序,作為蛋白質合成的直接模板。
⑵tRNA的功能是轉運胺基酸。在蛋白質合成過程中,tRNA與合成蛋白質所需的單體——胺基酸形成複合物,將胺基酸轉運到核糖體中mRNA的特定位置上。
與反轉錄
簡介
定義:以反義RNA為模版,通過反轉錄酶,進行的RNA轉錄
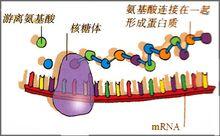 信使RNA
信使RNA1.概念反轉錄是以RNA為模板合成DNA的過程,也稱逆轉錄。這是DNA生物合成的一種特殊方式。
2.反轉錄酶與反轉錄過程
反轉錄過程由反轉錄酶催化,該酶也稱依賴RNA的DNA聚合酶(RDDP),即以RNA為模板催化DNA鏈的合成。合成的DNA鏈稱為與RNA互補DNA(cDNA)。反轉錄酶存在於一些RNA病毒中,可能與病毒的惡性轉化有關。人類免疫缺陷病毒(HIV)也是一種RNA病毒,含有反轉錄酶。在小鼠及人的正常細胞和胚胎細胞中也有反轉錄酶,推測可能與細胞分化和胚胎髮育有關。
生物學意義
反轉錄的發現有重要的理論意義和實踐意義。
1.對分子生物學的中心法則進行了修正和補充,修正後的中心法則表示為:
2.在致癌病毒的研究中發現了癌基因,在人類一些癌細胞如膀胱癌、小細胞肺癌等細胞中,也分離出與病毒癌基因相同的鹼基序列,稱為細胞癌基因或原癌基因。癌基因的發現為腫瘤發病機理的研究提供了很有前途的線索。
3.在實際工作中有助於基因工程的實施。由於目的基因的轉錄產物易於製備,可將mRNA反向轉錄形成DNA用以獲得目的基因。
可扼殺信使
microRNA雖然只有19-21個核苷長度,但能夠通過綁定和中和負責蛋白翻譯的信使RNA,沉默大片段基因。迄今發現的microRNA已達到幾百種,它們在基因組中的調節作用日益引起人們關注。
最近,Wistar研究所的研究人員發現microRNA會經歷一種生理學後果嚴重的分子編輯過程(molecular editing),序列的一個替換會改變這些microRNA的作用靶標,抑制非預期匹配物的表達,編輯過程出錯會導致嚴重的健康問題。詳細內容刊登於2月23日《Science》雜誌。
“我們發現,某些情況下已編輯的microRNA版本與未經編輯的版本只有一個核苷差異,”文章高級作者、Wistar研究所基因表達和校準程式教授 Kazuko Nishikura博士說,“這些已編輯的microRNA不是由DNA編碼的,意味著至少兩個版本來自於同一個基因,這是未曾料到的……進一步觀察,我們發現替換隻出現在分子的特定關鍵區域,總共19或21個核苷長度分子的第7或8位核苷,這兩個位點規定了靶標的特異性。提示替換可能會使已編輯的microRNA與未編輯的microRNA沉默完全不同的靶標。”
利用生物信息學工具將一種microRNA的已編輯版本和未編輯版本與已知的基因序列資料庫比對,研究人員鑑別出分別是兩種不同版本分子的靶標的兩組完全不同的基因,並從每組中分別挑選出三個基因,進一步觀察microRNA是否會改變這些基因的表達效果。
接下來,研究人員隨機挑選出一個預期的靶標基因,探測microRNA的衍生物的潛能。所挑選的基因為PRPS1,其所編碼的酶在尿酸合成中發揮關鍵作用。如果調節此酶不當,多種健康問題會接踵而來。比如,表達量過高會引起血液中尿酸水平上升,引起風濕痛;大腦中尿酸水平過高會損傷感覺神經元,引起失聰。
在不能進行RNA編輯(RNA editing)的轉基因小鼠和正常小鼠身上的實驗顯示,完全缺乏已編輯版本microRNA的小鼠,PRPS1酶水平是對照組的2倍,尿酸水平也是對照組的2倍。
Nishikura 說這些證實,最初計算機所預測的同一基因編碼的未經編輯的、已編輯的microRNA的不同靶標是正確的。至少對於PRPS1來說是正確的,未經編輯的和已編輯的microRNA的差異的生理學效果在尿酸水平的上升程度上顯示出來。
即便說PRPS1基因是研究人員隨機挑選的,但研究結果提示一系列其它迄今原因未明的疾病很有可能是由新發現的microRNA編輯過程引起的。
提取分離純化
真核細胞的mRNA分子最顯著的結構特徵是具有5’端帽子結構(m7G)和3’端的Poly(A)尾巴。絕大多數哺乳類動物細胞mRNA的3’端存在20-30個腺苷酸組成的Poly(A)尾,通常用Poly(A+)表示。這種結構為真核mRNA的提取 ,提供了極為方便的選擇性標誌,寡聚(dT)纖維素或寡聚(U)瓊脂糖親合層析分離純化mRNA的理論基礎就在於此。mRNA的分離方法較多,其中以寡聚(dT)-纖維素柱層析法最為有效,已成為常規方法。此法利用mRNA 3’末端含有Poly(A+)的特點,在RNA流經寡聚(dT)纖維素柱時,在高鹽緩衝液的作用下,mRNA被特異地結合在柱上,當逐漸降低鹽的濃度時或在低鹽溶液和蒸餾水的情況下,mRNA 被洗脫,經過兩次寡聚(dT)纖維柱後,即可得到較高純度的mRNA。寡聚(dT)纖維素柱純化mRNA
試劑準備
1.3M醋酸鈉(pH 5.2) 2.0.1M NaOH 3.1×上樣緩衝液:20mM Tris-HCl(pH 7.6);0.5M NaCl;1M EDTA(pH 8.0);0.1%SLS(十二烷基氨酸鈉。配製時可先配製Tris-HCl(pH 7.6)、NaCl、EDTA(pH 8.0)的母液,經高壓消毒後按各成分確切含量,經混合後再高壓消毒,冷卻至65℃時,加入經65℃溫育(30min)的10%SLS至終濃度為0.1%。4.洗脫緩衝液:10mM Tris-HCl(pH 7.6);1mM EDTA(pH 8.0);0.05% SDS 5.無水乙醇、70%乙醇 6.DEPC
操作步驟
1.將0.5-1.0g寡聚(dT)-纖維懸浮於0.1M的NaOH溶液中。2.用DEPC處理的1ml注射器或適當的吸管,將寡聚(dT)-纖維素裝柱0.5-1ml,用3倍柱床體積的DEPC H2O洗柱。3.使用1×上樣緩衝液洗柱,直至洗出液pH值小於8.0。4.將RNA溶解於DEPC H2O中,在65℃中溫育10min左右,冷卻至室溫後加入等體2×上樣緩衝液,混勻後上柱,立即收集流出液。當RNA上樣液全部進入柱床後,再用1×上樣緩衝液洗柱,繼續收集流出液。5.將所有流出液於65℃加熱5min,冷卻至室溫後再次上柱,收集流出液。6.用5-10倍柱床體積的1×上樣緩衝液洗柱,每管1ml分部收集,OD260測定RNA含量。前部分收集管中流出液的OD260值很高,其內含物為無Poly(A)尾的RNA。後部分收集管中流出液的OD260值很低或無吸收。7.用2-3倍柱容積的洗脫緩衝液洗脫Poly(A+)RNA,分部收集,每部分為1/3-1/2柱體積。8.OD260測定Poly(A+)RNA分布,合併含Poly(A+)RNA的收集管,加入1/10體積3M NaAc(pH5.2)、2.5倍體積的預冷無水乙醇,混勻,-20℃放置30min。9.4℃離心,10000g×15min,小心吸棄上清。用70%乙醇洗滌沉澱。[注意:此時Poly(A+)RNA的沉澱往往看不到]。4℃離心,10000g×5min,棄上清,室溫晾乾。10. 用適量的DEPC H2O溶解RNA。
注意事項
1.整個實驗過程必須防止Rnase的污染。2.步驟⑷中將RNA溶液置65℃中溫育然後冷卻至室溫再上樣的目的有兩個,一個是破壞RNA的二級結構,尤其是mRNA Poly(A+)尾處的二級結構,使Poly(A+)尾充分暴露,從而提高Poly(A+)RNA的回收率;另一個目的是能解離mRNA與rRNA的結合,否則會導致rRNA的污染。所以此步驟不能省略。3.十二烷基肌氨酸鈉鹽在18℃以下溶解度下降,會阻礙柱內液體流動,若室溫低於18℃最好用LiCl替代NaCl。4.寡聚(dT)-纖維素柱可在4℃貯存,反覆使用。每次使用前應該依次用NaOH、滅菌 ddH2O、上樣緩衝液洗柱。5.一般而言,107哺乳動物培養細胞能提取1-5μg Poly(A+)RNA,約相當於上柱總RNA量的1%-2%。
