
 轉譯
轉譯概述
蛋白質生物合成包括胺基酸的活化及其與專一轉移核糖核酸(tRNA)的連線;肽鏈的合成(包括起始、延伸和終止)和新生肽鏈加工成為成熟的蛋白質3大步驟。其中心環節是肽鏈的合成。蛋白質生物合成需核糖體、mRNA、tRNA、氨醯轉移核糖核酸(氨醯tRNA)合成酶、可溶性蛋白質因子等大約200多種生物大分子協同作用來完成。
 《DN完整版規則4》
《DN完整版規則4》胺基酸的活化及其與專一tRNA的連線
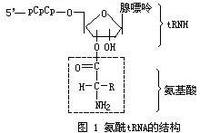 氨醯tRNA的結構
氨醯tRNA的結構氨醯tRNA的結構 胺基酸與tRNA反應的整個過程分兩步進行(見轉移核糖核酸),其總反應式表示如下:
胺基酸+ATP+tRNA氨醯tRNA+AMP+Ppi
上述反應都是在氨醯tRNA合酶催化下進行的。此酶具有高度專一性,每種胺基酸至少有一種氨醯tRNA合成酶。不同氨醯tRNA合成酶在大小、亞基結構和胺基酸組成上各不相同,其分子量大多在85000~110000之間,其中有些酶已製得結晶。
肽鏈的合成
肽鏈的合成分3個步驟:起始、延伸、終止。合成方向從氨基端(N端)向羧基端(C端)進行。mRNA的翻譯方向則是從5′端→3′端。
1、起始
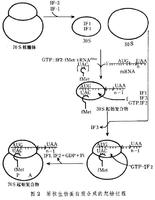 原核生物蛋白質合成的起始過程
原核生物蛋白質合成的起始過程原核生物蛋白質生物合成的起始因子有3種——IF-1、IF-2和IF-3,參與起始的氨醯tRNA(也叫起始tRNA)是甲醯甲硫氨醯tRNA(fMet-tRN原核生物蛋白質合成的起始過程AfMet),其中甲醯基是在甲醯化酶催化下加到甲硫氨醯tRNA上的。起始過程分以下3步:①70S核糖體在起始因子IF-3和IF-1作用下解離,產生30S和50S兩個亞基。②30S亞基與mRNA起始密碼子部位結合,fMet-tRNAfMet在IF-2作用下,並有GTP參與,進入30S亞基,釋放出IF-3,形成30S起始複合物。在這個複合物中,fMet-tRNAfMet上的反密碼子與mRNA上的起始密碼子(翻譯開始的信號)之間形成互補鹼基對。③30S起始複合物與50S亞基結合,IF-2(具有依賴於核糖體的GTP水解酶活性)水解GTP,產生GDP和無機磷,並釋放出能量,使IF-2,IF-1和GDP等從複合物中釋放出來,形成70S起始複合物(包括70S核糖體、mRNA和fMet-tRNAfMet)。這時,fMet-tRNAfMet占據核糖體上的肽基-tRNA位置(P位)。70S起始複合物巳具備了肽鏈延伸的條件(圖2)。
真核生物肽鏈合成的起始因子比原核的多(如兔網織細胞至少有9種),起始tRNA是甲硫氨醯tRNA(Met-tRNAMet),不同於原核生物的fMet-tRNAfMet。起始基本步驟與原核生物的相同,也包括核糖體的解離,小亞基(40S)起始複合物的形成和肽鏈起始複合物(80S)的形成。主要區別在於真核生物的核糖體小亞基先與氨醯化的起始tRNA結合,然後再與mRNA結合;而原核生物核糖體小亞基在形成起始複合物時則先與mRNA結合,再與起始tRNA結合。
2、延伸
肽鏈合成的循環過程
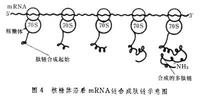 核糖體沿著mRNA鏈合成肽鏈示意圖
核糖體沿著mRNA鏈合成肽鏈示意圖在肽鏈延伸過程中,當第1個核糖體沿mRNA移動到離起始密碼子較遠(約40個核苷酸)時,第2個核糖體又與起始密碼子結合併開始另一條新肽鏈的合成,同樣第3、第4個核糖體相繼與同一mRNA結合,從而形成多核糖體。體內蛋白質合成實際上是以多核糖體的形式進行的(圖4)。肽鏈合成的終止
3、終止
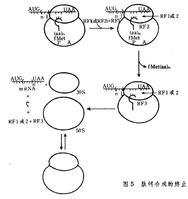 肽鏈合成的終止
肽鏈合成的終止新生肽鏈(蛋白質前體)的加工
 核糖體進行轉譯
核糖體進行轉譯蛋白質生物合成的調控
生物體內蛋白質合成的速度,主要在轉錄水平上,其次在翻譯過程中進行調節控制。它受性別、激素、細胞周期、生長發育、健康狀況和生存環境等多種因素及參與蛋白質合成的眾多的生化物質變化的影響。由於原核生物的翻譯與轉錄通常是偶聯在一起的,且其mRNA的壽命短,因而蛋白質合成的速度主要由轉錄的速度決定。弱化作用是一個通過翻譯產物的過量與不足首先影響轉錄,從而調節翻譯速度的一種方式。mRNA的結構和性質也能調節蛋白質合成的速度。
真核起始因子對蛋白質生物合成的調控作用
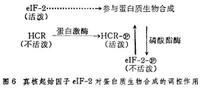 真核起始因子對蛋白質生物合成的調控作用
真核起始因子對蛋白質生物合成的調控作用血紅素通過影響eIF-2對蛋白質進行調控。當血紅素存在時,抑制了細胞蛋白質合成,而且還能促進通常不合成血紅蛋白的細胞合成蛋白質,如促進肝癌細胞、海拉細胞和腹水瘤細胞無細胞製劑的蛋白質合成。
蛋白質生物合成的抑制劑
蛋白質生物合成抑制劑
 蛋白質生物合成抑制劑
蛋白質生物合成抑制劑