
物種進化
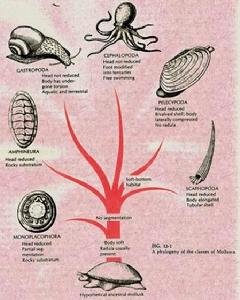
軟體動物的海產種類個體發生中為螺鏇型卵裂,且具有擔輪幼蟲,排泄器官為後腎管,這些特點均與環節動物尤其是多毛類近似。故有理由認為軟體動物和環節動物在系統發生中有著共同的起源,在長期進化中,朝著不活動的生活方式發展,因而體節消失,產生了貝殼,運動器官和神經感官均趨於退化。軟體動物中單板綱、無板綱及多板綱較為原始,這幾類的次生體腔發達,近似梯式神經;有的體呈蠕蟲形,無殼,許多器官如鰓、腎、外殼等顯不出分節排列現象。這些原始性狀的存在認為它們接近軟體動物的原始祖先,各自獨立發展一支。
腹足類較為原始,其生活方式活躍,頭部發達。瓣鰓綱生活方式不活動無頭,但原始種類具盾鰓,足部具趾面,這與腹足綱接近。掘足綱頭不明顯,套膜在胚胎時為2片,後才癒合呈筒狀,成對的腎,腦神經節與側神經節分開,這些表明接近於原始的瓣鰓類、但掘足類無鰓,無心臟,貝殼筒形,又顯示與其他綱動物在演化上較為疏遠,可能是較早分出的一支。頭足綱為一古老的類群,起源早,化石種類多。它們生殖腔與體腔相通,似無板綱;個體發生中在胚胎早期無腎,似多板綱和無板綱;生殖導管來源於體腔導管又似多板綱。由於頭足綱其有原始軟體動物的特點說明它們與軟體動物的原始種類接近。但頭足綱有機結構,複雜神經系統高度集中,且為軟骨質包圍;眼的結構似脊椎動物,基本為閉管式循環系統;直接發生,無幼蟲期。由於頭足綱既有原始性狀,又有高度的進化特徵,故推測它們可能很早分出的一支,沿著更為活躍的生活方式發展的一個獨立的分支。
外形特徵
頭部
位於身體前端。一些行動遲緩的原始種類頭部不發達,僅有口,與身體沒有明顯的界限,如石鱉等;一些穴居或固著生活的種類體軀完全包被於外套膜和貝殼之內,頭部退化,如蚌類、牡蠣等;一些比較進化、運動敏捷的種類頭部發達,分化明顯,生有觸角和眼等感覺器官,如田螺、蝸牛及烏賊等。
足部
足部是位於身體腹側的運動器官,隨生活方式不同呈現不同形式:有的種類足部蹠面平滑,適於在陸地或水底爬行,如腹足綱;有的種類足部呈斧刃狀,有利於挖掘泥沙,如瓣鰓綱;有些固著生活的種類足退化,如牡蠣科;也有些種類足部萎縮,失去了運動功能,但有足絲腺,能分泌足絲,用以附著在外物上生活,如貽貝科、扇貝科等。在頭足綱,足生於頭部,有的特化成腕,上面生有許多吸盤,為捕食器官,並有一部分變態成漏斗,適於游泳生活,如烏賊和章魚等。少數種類足的側部(即側足,parapodium)特化成片狀,可游泳,稱為翼或鰭,如翼足目(Pteropoda)。足部通常生有平衡器,有些種類在足的上部生有許多觸手。
生長繁殖
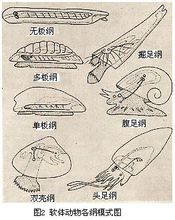 軟體動物門
軟體動物門軟體動物的生殖系統由生殖腺、生殖輸送管、交接器和一
些附屬腺體構成。生殖腺由體腔壁形成。生殖輸送管內端通向生殖腺腔,外端開口於外套腔或直接開口於體外。軟體動物有雌雄異體和雌雄同體之分。雌雄異體的種類包括多板綱、絕大多數的前鰓亞綱和瓣鰓綱、頭足綱等,它們有的通過交尾受精,有的將生殖產物分別排到水中受精。雌雄同體的種類包括無板綱、後鰓亞綱、肺螺亞綱以及少數的前鰓類和瓣鰓綱,它們大多通過交尾受精。
軟體動物的受精卵是典型的螺鏇型卵裂,由外包或內陷或由二者形成原腸胚,原腸胚形成後,很快發育為自由游泳的擔輪幼蟲。個別種類從擔輪幼蟲直接發育成成體,但大多數種類從擔輪幼蟲發育成面盤幼蟲(veliger larva),然後才發育成成體。擔輪幼蟲的形態與環節動物多毛類的幼蟲近似,面盤幼蟲發育早期背側有外套的原基,且分泌外殼,腹側有足的原基,口前纖毛環發育成緣膜(velum)或稱面盤。大多數的海產腹足類的擔輪幼蟲在卵袋中度過,一些前鰓類和淡水腹足類、肺螺類的擔輪幼蟲和面盤幼蟲都在卵袋中度過。在淡水中生活的蚌類,面盤幼蟲特化為適應寄生生活的鉤介幼蟲(glochidium),這種幼蟲在魚類的鰓、鰭或其他部位寄生,在魚體上形成胞囊。幼蟲從寄主身體獲取營養,逐漸發育成成體,破囊而出,沉落水底營底棲生活。頭足綱的卵子分裂屬子不完全分裂的盤狀分裂類型,為直接發育。
棲息環境
腹足類在陸地、淡水(湖與小溪)和海洋均有分布,瓣鰓綱只生活在淡水和海洋中,其他類群則完全生活在海洋中。
引言
軟體動物的數量僅次於節肢動物,約有10萬種,為動物界中第二大類群,包括各種螺類、蚌類、烏賊、章魚等。
軟體動物雖然在外形上差別很大,但主要形態結構基本相同。內臟結構較環節動物複雜,但在進化特徵上沒有明顯的進步。從胚胎髮育角度分析,軟體動物也經過擔輪幼蟲期,說明軟體動物和環節動物兩者親緣關係極為密切。軟體動物經濟意義較大,絕大多數種類可供食用。有的軟體動物可以作藥用,如鮑魚的貝殼是中藥石決明,烏賊的內骨骼是海螵蛸。珍珠既可入藥,又是名貴裝飾品。有些軟體動物對人類有害,鑿船貝(teredo)嚴重危害海中木船和木質建築,蝸牛是田園、果樹和農作物害蟲,釘螺等是寄生蟲的中間寄主。
概況
 軟體動物門
軟體動物門(Mollusca)物種類繁多,生活範圍極廣,海水、淡水和陸地均有產。已記載130000多種,僅次於節肢動物,為動物界的第二大門。軟體動物的結構進一步複雜,機能更趨於完善,它們具有一些與環節動物相同的特徵:次生體腔,後腎管,螺鏇式卵裂,個體發育中具有擔輪幼蟲等,因此認為軟體動物是由環節動物演化而來,朝著不很活動的生活方式較早分化出來的一支。
本門動物體外大都復蓋有各式各樣的貝殼,故通常又稱之為貝類。由於它們大多數貝殼華麗,肉質鮮美,營養豐富,又較易捕獲,因此遠在上古漁獵時期,就已被人類利用。其中不少可供食用、藥用、農業用、工藝美術業用,也有一些種類有毒,能傳播疾病,危害農作物,損壞港灣建築及交通運輸設施,對人類有害。
主要特徵
軟體動物的形態結構變異較大,但基本結構是相同的。身體柔軟,不分節,可區分為頭、足、內臟團三部分,體外被套膜,常常分泌有貝殼。
◆身體的劃分
軟體動物的身體一般可分為頭、足和內臟團三個部分。
①頭部
位身體的前端。運動敏捷的種類,頭部分化明顯,其上生有眼、觸角等感覺器官,如田螺、蝸牛及烏賊等;行動遲緩的種類頭部不發達,如石鱉;穴居或固著生活的種類,頭部已消失,如蚌類、牡蠣等。
②足部
通常位於身體的腹側,為運動器官,常因動物的生活方式不同,而形態各異。有的足部發達呈葉狀、斧狀或柱狀,可爬行或掘泥沙;有的足部退化,失去了運動功能,如扇貝等;固著生活的種類,則無足,如牡蠣;有的足已特化成腕,生於頭部,為捕食器官,如烏賊和章魚等,稱為頭足;少數種類足的側部(即側足parapodium)特化成片狀,可游泳,稱為翼或鰭,如翼足類(Pteropoda)。
③內臟團(visceral mass)
為內臟器官所在部分,常位於足的背側。多數種類的內臟因為左右對稱,但有的扭曲成螺鏇狀,失去了對稱形,如螺類。
◆外套膜(mantle)
為身體背側皮膚褶向下伸展而成,常包裹整個內臟團。外套膜與內臟團之間形成的腔稱外套腔(mantle cavity)。腔內常有鰓、足以及肛門、腎孔、生植孔等開口於外套腔。
外套膜由內外兩層上皮構成,外層上皮的分泌物,能形成貝殼,內層上皮細胞具纖毛,纖毛擺動,造成水流,使水循環於外套腔內,藉以完成呼吸、排泄、攝食等。左右2片套膜在後緣處常有一二處癒合,形成出水孔(exhalant siphon)和入水孔(inhalant siphon)。有的種類出入水孔延長成管狀,伸出殼外稱為出水管和入水管。
體外具貝殼為軟體動物的重要特徵,因此研究軟體動物的學科又稱貝類學(Malacology)。大多數軟體動物都具有一二個或多個貝殼,形態各不相同。有的呈帽狀;螺類為螺鏇形;掘足類為管狀;瓣鰓類為瓣狀。有些種類的貝殼退化成內殼,有的無殼。貝殼有保護柔軟身體的功能。
貝殼的成分主要是碳酸鈣和少量的殼基質(conchiolin或稱貝殼素)構成,這些物質是由外套膜上皮細胞分泌形成的。貝殼的結構一般可分為3層),最外一層為角質層(periostracum),很薄,透明,有光澤,由殼基質構成,不受酸鹼的侵蝕,可保護貝殼。中間一層為殼層(ostracum),又稱稜柱層(primatic layer)占貝殼的大部分,由角狀的方解石(cacalcite)構成。最內一層為殼底(hypostracum),即珍珠質層(peral layer),富光澤,由葉狀霰石(aragonite)構成。外層和中層為外套膜邊緣分泌形成,可隨動物的生長逐漸加大,但不增厚;內層為整個套膜分泌而成,可隨個體的生長而增加厚度。珍珠就是由珍珠質層形成的。當外套膜受到微小砂粒等異物侵人刺激,受刺激處的上皮細胞即以異物為核,陷入外套膜的上皮之間結締組織中,陷入的上皮細胞自行分裂形成珍珠囊,囊即分泌珍珠質,層復一層地將核包位逐漸形成珍珠。據史料記載公元前2200多年,中國就有淡水育珠的記載(書經.禹貢篇),廣西合浦育珠自古就很有名,採摘開始於漢代。
角質層和稜柱層的生長非連續不斷的,由於食物、溫度度等因素影響外套膜分泌機能,故貝殼的生長速度是不同的,因此在貝殼表面形成了生長線,表示出生長的快慢。
器官
◆消化系統
軟體動物的消化管發達,少數寄生種類(內寄螺Entocolax)退化。多數種類口腔內具顎片(mandible)和齒舌(radula),顎片一個或成對,可輔助捕食。齒舌是軟體動物特有的器官,位口腔底部的舌突起(odontophore)表面,由橫列的角質齒組成,似挫刀狀。攝食時以齒舌作前後伸縮運動刮取食物。齒舌上小齒的形狀和數目,在不同種類間各異,為鑑定種類的重要特徵之一。小齒組成橫排,許多排小齒構成齒舌。每一橫排有中央齒一個,左右側齒一或數對,邊緣有緣齒一對或多對。齒舌上小齒的排列以齒式表示如中國圓田螺(Cipangopaludina chinensis)的齒式為2.1.1.1.2。
軟體動物的次生體腔極度退化,殘留圍心腔(pericardinal cavity)及生殖腺和排泄器官的內腔。初生體腔則存在於各組織器官的間隙,內有血液流動,形成血竇(blood sinus)。
循環系統由心臟、血管、血竇及血液組成。心臟一般位內臟團背側圍心腔內,由心耳和心室構成。心室一個,壁厚,能博動,為血循環的動力;心耳一個或成對,常與鰓的數目一致。心耳與心室間有辯膜,防止血液逆流。血管分化為動脈和靜脈。血液自心室經動脈,進人身體各部分,後匯入血竇,由靜脈回到心耳,故軟體動物為開管式循環。一些快速游泳的種類,則為閉管式循環。血液無色,內含有變形蟲狀細胞。有些種類血漿中含有血紅蛋白(haemoglobin)或血青蛋白(haemocyanin),故血液呈紅色或青色。
◆呼吸器官
水生種類用鰓呼吸,鰓為外套腔內面的上皮伸展形成,位腔內。鰓的形態各異,鰓軸兩側均生有鰓絲,呈羽狀,稱盾鰓;僅鰓軸一側生有鰓絲,呈梳狀,稱櫛鰓(ctenidium);有的鰓成瓣狀,稱瓣鰓(lamellibranch);有些種類的鰓延長成絲狀。稱絲鰓(filibranch)。有的本鰓消失,又在背側皮膚表面生出次生鰓(secondary branchium),也有的種類無鰓。鰓成對或為單個,數目不一,少則一個或一對,多則可達幾十對。陸地生活的種類均無鰓。其外套腔內部一定區域的微細血管密集成同形成肺,可直接攝取空氣中的氧。這是對陸地生活的一種適應性。
◆排泄器官
軟體動物的排泄器官基本上是後腎管,其?原腎管。後腎管由腺質部分和管狀部分組成,腺質部分富血管,腎口具纖毛,開口於圍心腔;管狀部分為薄壁的管子,內壁具纖毛,腎孔開口於外套腔。後腎管不僅可排除因心臟中的代謝產物,也可排除血液中的代謝產物。另外圍心臟內壁上的圍心臟腺,微血管密布,可排除代謝產物於圍心臟內,由後腎管排出體外。
原始種類的神經系統無神經節的分化,僅有圍咽神經環及向體後伸出的一對足神經索(pedal cord)和一對側神經索(pleural cord)。較高等的種類主要有 4對神經節,各神經節間有神經相連。腦神經節(cerebral ganglion)位食管背側,發出神經至頭部及體前部,司感覺;足神經節(pedal ganglion位足的前部,伸出神經至足部,司運動和感覺;側神經節(peural ganglion)發出神經至外套膜及鰓等;髒神經節(visceral ganglion)發出神經至各內臟器官。這些神經節有趨於集中之勢,有的種類的主要神經節集中在一起形成腦,外有軟骨包圍,如頭足類。軟體動物已分化出觸角、眼、嗅檢器及平衡囊等感覺器官,感覺靈敏。
軟體動物大多數為雌雄異體,不少種類雌雄異形;也有一些為雌雄同體。卵裂形式多為完全不均等卵裂,許多屬螺鏇型。少數為不完全卵裂。個體發育中經擔輪幼蟲和面盤幼蟲(veliger larva)兩期幼蟲,擔輪幼蟲的形態與環節動物多毛類的幼蟲近似,面盤幼蟲發育早期背側有外套的原基,且分泌外殼,腹側有足的原基,口前纖毛環發育成緣膜(velum)或稱面盤。也有的種類為直接發育。淡水蚌類有特殊的鉤介幼蟲(glochidium)。
分類
依據它們的貝殼、足、鰓、神經及發生特點等特徵,分為七個綱。
◆單板綱(Monoplacomphora)
絕大多數為化石種,已絕滅了近4億年。這類動物有一個帽狀或匙形的貝殼,有2~8對對稱的肌痕。1952年丹麥海洋研究船在太平洋沿岸哥斯大黎加3570m深海處第一次採得生活標本,1957年被定名為新碟貝(Neopilina galathea lemche),以後又在大西洋、印度洋等處發現了一些種類。
新碟貝是一類原始的貝類,體為兩側對稱,具一近圓形而扁的貝殼,腹足強大,周緣肌肉發達,中央簿,故無吸附能力,僅適於在海底滑行。縮足肌8對,分節排列於足的周圍。足四周為外套溝,兩側共有鰓5或6對。足前端為口,後端為肛門。口前有一對具纖毛的口蓋(velum),口後有扇狀觸手一對。心臟位圍心腔內,由一心室及2對心耳構成;腎6或7對;雌雄異體,生殖腺2對,有生殖導管開口於腎,生殖細胞由腎排出體外。神經系統由圍食道神經環及向後伸出的側神經和足神經組成。
這類被稱為活化石的原始貝類的發現,對研究貝類的起源與演化提供了新的資料。
◆無板綱(Aplacophora)
為軟體動物中的原始種類,體呈蠕蟲狀.細長或短粗,無貝殼。體表被具石灰質細棘的角質外皮,頭小,口在前端腹側,軀體細長,腹側中央有一腹溝,有的種類溝中有一小形其纖毛的足,有運動功能.體後有排泄腔,多數種類在腔內有一對鰓,腔後為肛門、無板類無觸角、眼等感覺器官:腸為直管狀,齒舌有或無;心臟為一心室一心耳,血管系統退化,雌雄同體或異體,個體發生中有擔輪幼蟲期。
無板類約有200多種,生活在低潮線下數10m至深海海底,肉食性分布遍及全球。我國南海海域79m深處曾採得龍女簪一種。
◆多板綱(Polyplacophora)
全部生活在沿海潮間帶,常以足吸附於岩石或藻類上。體呈橢圓形,背稍隆,腹平。背側具八塊石灰質貝殼,多是復瓦狀排列、前面一塊半月形,稱頭板(cephalic plate),中間 6塊結構一致、稱中間板(intermediate plate),未塊為元寶狀,稱尾板(tail plate)。各板間可前後抽拉移動,因此動物脫離岩石後,可以曲捲起來。貝殼周圍有一圈外套膜,稱環帶(girdle), 其上叢生有小針、小棘等,形態各異、頭部不發達,位腹側前方,圓柱狀,有一向下的短吻,吻中央為口。足寬大,吸附力強,在岩石表面可緩慢爬行。足四周與外套之間有一狹溝,即外套溝,在溝的兩側各有一列盾鰓,六對或數十對。
多板類約有1000種,中國沿海習貝種類有毛膚石鱉體型較大,環帶上有成叢的白色針束;鱗帶石鱉殼片高,頭板具放射肋。
軟體動物中最大的一類,有10萬種以上。生活在海洋、淡水及陸地,分布遍及全球,少數種類為寄生(內寄螺Entocolax、光螺Melanella、內殼螺Entoconcha等)。
◆掘足綱(Scaphopoda)
全海產。具長圓錐形稍彎曲的管狀貝殼,如象牙狀。粗的一端為前端,開口大。稱為頭足孔;細的一端為後端,開口小,稱為肛門孔。殼凹的一面為背側,凸的一面為腹側。外套膜呈管狀,前後端有開口。頭部不明顯,前端具有不能伸縮的吻,吻基部兩側生有許多頭絲(captacula),能伸縮,末端膨大。頭絲可伸出殼外,有觸覺功能,也可攝食。掘足類為肉食性,吻內為口球,具顎片和齒舌。足在吻的基部之後,柱狀,末端三葉狀或盤狀。足可伸得很長,能挖掘泥沙。肛門開口於足的基部腹側。無鰓,以外套膜進行氣體交換。循環器官心臟一室無心耳,未分化出血管,僅有血竇。腎一對,囊狀,位胃側面。雌雄異體,生殖腺一個;個體發生中有擔輪幼蟲和面盤幼蟲。
掘足類自潮間帶至4000m深海都有分布,約300種僅2科。角貝殼呈象牙狀,足圓錐形,末端有二冀狀側葉。本綱動物在我國分布廣,種類多。
◆瓣鰓綱(Lamellibranchia)
瓣鰓綱動物全部生活在水中,大部分海產,少數在淡水,極少數為寄生(內寄蛤Entovalva、戀蛤Peregrinamor等)。約有2萬種,分布很廣。一般運動緩慢,有的潛居泥沙中,有的固著生活,也有的鑿石或鑿木而棲。
◆頭足綱(Cephalopoda)
頭足類全海產,肉食性。體左右對稱,分頭、足、軀幹三部分。頭部發達,兩側有一對發達的眼,原始種類具外殼,多數為內殼或無殼;足著生於頭部,特化成腕和漏斗,故稱頭足類。漏斗位於頭腹面、頭與軀幹之間。羽狀鰓一對或2對,心耳和腎的數目與鰓一致。具軟骨。口腔有顎片和齒舌。神經系統集中,感官發達。閉管式循環系統。直接發生。
頭足類的生活種類約有700種化石種類在10000種以上。
生活習性
1、浮游生活:營這種生活的種類都是隨波逐流地在海洋中過漂浮生活。一般個體較小,貝殼很薄或沒有貝殼。有的種類足特化成鰭,如翼足類和異足類中的許多種。有的種類由足分泌一個浮囊攜帶動物在海洋中漂浮,如海蝸牛。由於它們是隨波逐流的,所以分布範圍與海流有密切關係,例如中國近海浮游軟體動物的分布受台灣暖流的影響很大,暖流勢強時分布靠北,暖流勢弱時分布靠南。
2、游泳生活:營游泳生活的種類能和魚類一樣在海洋中長距離洄游,例如頭足綱的烏賊、槍烏賊、章魚等,它們的足特化成腕和漏斗,胴部兩側產生鰭,靠漏斗噴水和鰭的作用可以迅速平穩地遊動。某些瓣鰓綱(如扇貝、日月貝、銼蛤等)雖然不過游泳生活,但在必要時,可憑藉貝殼的急劇開閉和外套觸手的作用在海中作蝶式游泳。
3、底棲生活:絕大多數的軟體動物營底棲生活。它們在水底匍匐爬行,或在底質上固著。有的種類營底上生活,例如鮑、蜒螺、田螺、織紋螺、紅螺等在岩石或泥沙灘表面爬行;貽貝、扇貝等用足絲附著在海底岩石或其他外物上;牡蠣、猿頭蛤、海菊蛤等用貝殼固著在海底外物上。有的種類營底內生活,例如很多瓣鰓綱(蛤、蛤蜊、烏蛤、攖蛤、竹蟶等)靠發達的足部挖掘泥沙,把身體整個埋於底內棲息,靠水管與底表溝通。許多穿孔種類在木材、岩石、貝殼、珊瑚礁等堅硬的底質中生活,例如石蟶、海筍、開腹蛤穿鑿岩石、貝殼等,馬特海筍和船蛆穿鑿木材。
4、寄生生活:有的為外寄生,如圓柱螺寄生於棘皮動物腕的步帶溝中;有的為內寄生,如內殼螺寄生在錨海參的食道內。
經濟意義
軟體動物中有很多種類可以為人類所利用,有益於人類,但也有許多種類對危害人類常造成經濟上的損失。
與人類的關係
價值
①食用價值
海產的鮑、玉螺、香螺、紅螺、東風螺、泥螺、蚶、貽貝、扇貝、江珧、牡蠣、文蛤、蛤仔、蛤蜊、蟶、烏賊、槍烏賊、章魚,淡水產的田螺、螺螄、蚌、蜆,陸地棲息的蝸牛等肉味鮮美,具有很高的營養價值。
②藥用價值
鮑的貝殼(中藥稱石決明),寶貝的貝殼叫海巴,珍珠、烏賊的貝殼叫海螵蛸,以及蚶、牡蠣、文蛤、青蛤等的貝殼等都是中藥的常用藥材。從鮑魚、鳳螺、海蝸牛、蛤、牡蠣、烏賊等可以提取抗生素和抗腫瘤藥物。
③農業價值
產量多的小型軟體動物可以做農田肥料或飼料,河蜆可以飼養淡水魚類。④工業用。軟體動物的貝殼是燒石灰的良好原料。珍珠層較厚的貝殼( 如蚌、馬蹄螺等 )是制鈕扣的原料 。
④工藝用或裝飾
很多貝類的貝殼有獨特的形狀和花紋,富有光澤,絢麗多彩,是古今中外人士喜歡蒐集的玩賞品。
⑤地質價值
軟體動物門在地質歷史時期中有很多可作為指示沉積環境的指相化石。在世界和中國寒武系的最底部,已有單板綱和其他軟體動物化石出現,中生界的不少菊石成為洲際範圍內劃分、對比地層的帶化石,有些可用以了解古水域溫度和含鹽度等;蝸牛化石能反映第四紀氣候環境。
對人類的危害
陸生的蝸牛、蛞蝓等吃植物的葉、芽,危害蔬菜、果樹、菸草等;海洋中的一些肉食性種類,能殺害牡蠣、泥蚶等的幼苗,造成養殖雙殼類的損失;一些草食性種類常吃海帶、紫菜的幼苗,是藻類養殖的敵害。在淡水和陸生的軟體動物中,椎實螺是肝片吸蟲的中間宿主,豆螺是華枝睪吸蟲的中間宿主,扁卷螺是薑片蟲的中間宿主,短溝蜷是肺吸蟲的中間宿主,釘螺是日本血吸蟲的中間宿主,對人類的危害十分嚴重。海洋中的船蛆、海筍等是專門穿鑿木材或岩石穴居的種類,對於海洋中的木船、木樁和海港的木、石建築都有危害。營附著或固著生活的種類常大量附著在船底,可以影響船隻的航行速度。有些附著生活的種類,可以堵塞水管,影響生產。
