
簡介

漸新世
漸新世是第三紀的第三個世,約開始於3650萬年前,結束於2330萬年前。漸新世時形成的地層稱漸新統 ,位於始新統之上、中新統之下。
1833年,英國C.萊伊爾把第三紀分成始新世 、中新世和上新世.1854年,E.von 貝利希在德國發現早於中新世、晚於始新世的沉積物,從而提出漸新世。當時特提斯海周圍熱帶植物繁茂,德國的大部分地區溫暖潮濕,因此 ,形成大規模的褐煤。在漸新世時海生動物有孔蟲和貨幣蟲占優勢,陸生無脊椎動物豐富多樣。在波羅的海區有許多昆蟲,如蝴蝶、蜜蜂、螞蟻和蜘蛛。陸生脊椎動物也豐富多樣,分布於北美、歐洲、非洲和亞洲。
比起其他比較古老的地質時期,用岩床來確認漸新世是良好的識別方式,雖然精確的起始與結束時間有些不確定。漸新世(Oligocene)之名,詞源來自希臘文字ὀλίγος(oligos,“少”之意)和καινός(kainos,“新”之意),即表示在始新世噴髮式的進化之後,這個時期哺乳動物種類增加並不明顯。
漸新世被認為是一個重要的過渡時期,是一條連線“炎熱的始新世時期的古老世界和生態系統更具有現代特徵的中新世”的紐帶。漸新世時期生態系統發生的一個重要改變是草原在全球的擴張,而熱帶闊葉林則萎縮至赤道一帶。
標誌漸新世開始的事件是一起被稱為大置換的大規模物種滅絕事件,當時歐洲的動物群,除了區域性的嚙齒類動物和有袋類動物之外,都被來自亞洲的動物群所取代。漸新世和中新世則沒有顯著的全球性事件為其分野,而是各個地區相繼從較溫暖的漸新世晚期(2,600-2,300萬年前)進入較寒冷的中新世。
時期劃分
漸新世分為兩個時期:
晚漸新世(Chattian) (28.4 ± 0.1 – 23.03 百萬年)
早漸新世(Rupelian) (33.9 ± 0.1 – 28.4 ± 0.1 百萬年)
前一世 地質時代 後一世
始新世← 漸新世 →中新世
古地理
在這段時期,各大洲繼續飄移向它們目前所在的位置。南極洲越來越孤立,並最終成為一個永久性的冰封大陸。
北美洲西部的造山運動仍在繼續;非洲板塊持續向北擠壓歐亞板塊,阿爾卑斯山脈隨之開始隆起,並將特提斯洋的殘餘部分與大洋隔離開。在漸新世早期,歐洲遭遇到了短期的海洋入侵,這種狀況在北美洲則較少發生。漸新世早期在北美洲和歐洲之間似乎仍有陸橋相連,因為這段時期這兩處的動物區系仍然十分相似。
在漸新世的某段時間裡,南美洲最終與南極洲分離,並漂向北美洲;這也使得南極洲環流得以暢通無阻,最終使南極大陸的溫度急劇下降。
氣候
古近紀時期全球氣溫持續下降的趨勢在漸新世被打斷,在40萬年之間氣溫急劇下降8.2℃之後進入了一段長達700萬年的氣溫平穩震盪期。這段平穩震盪期從3,250萬年前開始,至2,550萬年前結束。漸新世的氣候變遷包括全球冰量的增加、海平面的下降(降幅達55米,從3,570萬年前至3,350萬年前)和氣溫的平穩變化。之後,在2,800萬年至2,600萬年間,隨著LaGaritaCaldera超級火山的爆發,這段氣溫平穩震盪期隨之結束。不過,漸新世和中新世更替期間的氣候仍然比較穩定。
植被
被子植物繼續在全球擴張;熱帶和亞熱帶森林則被溫帶落葉林所取代。開闊的平原和沙漠面積較之原來擴大了許多。草原也從始新世時期的河岸地帶發展至開闊地帶,但仍未發展至今日熱帶和亞熱帶大草原之規模。
在北美洲,亞熱帶物種,如今日的荔枝和腰果等,占據著主導地位;而山毛櫸、薔薇屬和松屬植物亦十分常見。帶有豆莢的豆科植物、莎草科植物和蕨類植物都繼續繁衍興盛。
動物種類
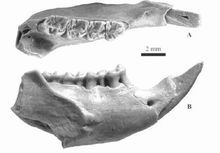 漸新世生物
漸新世生物漸新世時期,南美洲與其他大陸相互隔離,在其上發展出了十分特別的動物區系,南美洲成為了許多奇異動物,如焦獸目、閃獸目、滑距骨目和南方有蹄目動物的繁衍地。Sebecosuchian鱷魚、駭鳥和肉食性有袋類動物,如袋鬣狗科動物,仍是占支配地位的捕食者。雷獸在漸新世最初階段就已經滅絕了;而到了漸新世結束之時,裂齒類動物則在非洲和中東之外的地區消失了。多瘤齒獸目動物,作為一個古老的原始哺乳動物譜系,也在漸新世滅亡了。漸
 鬣齒獸
鬣齒獸漸新世的海洋動物與現今的物種十分相似,如雙殼綱動物。不過這個時期發現的海洋動物化石沒有始新世和中新世豐富。鬚鯨和齒鯨已經在這個時期出現,而他們的始祖古鯨亞目動物由於缺乏回聲定位能力(這種能力在當時海水變冷和變渾濁的情況下十分有用),種群開始衰退,造成其消亡的原因還包括氣候變化、來自現代鯨類和亦在該時期出現的頁鯊的競爭。早期的鏈齒獸目動物,如河馬眼索齒獸(Behemotops),其化石最早即出現在漸新世。在漸新世末期,由其與熊和水獺近似的始祖進化而來的鰭足類動物可能出現了。
三達河組
 漸新世
漸新世三達河組的沉積岩層現被分成兩個岩性段:上部早漸新世晚期—晚漸新世山地段(Shand Member)和下部早漸新世早期塔塔爾段(Tatal Member);兩岩段間有時夾有Khar Khoroo玄武岩層(Bryant and McKenna,1995;Meng and McKenna,1998)。該玄武岩層的時代用40Ar/39Ar法被確定為距今31。5 Ma(Hock et al。,1999,他們稱其為Basalt Ⅰ)。
三達河組中產有豐富的哺乳動物化石,特別是嚙齒類化石(Matthew and Granger,1923;Kowalski,1974;Russelland Zhai,1987;Hock et al。,1999)。在三達河組中採集到一批嚙齒類化石,其中包括松鼠和山河狸2科的化石。
松鼠
現生的松鼠科分異很大,而且大多數都分布在亞洲。然而,松鼠科化石在古近紀時在北美和歐洲卻很豐富。在北美出現於晚始新世,在漸新世時已明顯分異;在歐洲出現於早漸新世,在中新世時分異很大。相反,亞洲古近紀的松鼠化石卻發現得很少,標本也很破碎。已報導的只有3件標本。2件產自黨河下游地區:Bohlin(1946)報導的晚漸新世的Sciurus sp。,標本只是2枚單個臼齒(M3和m2);王伴月、邱占祥(2004)報導的早漸新世的黨河鮮松鼠(Oligo-sciurus dangheensis),標本只是一段帶m1~2的下頜骨。還有一件是Minin(2004)報導的蒙古漸新世的三達河克熱姆鼠(Kherem hsandgoliensis),標本也只是一段帶m1~3的下頜骨。
松鼠化石產自蒙古國的巴彥洪戈爾省的塔石蓋貝(Tashgain Bel)地點的三達河組上部山地段,時代為早漸新世晚期—晚漸新世。這批松鼠化石標本包括1件具P3~M3的上頜骨(PC 36-9)和2段下頜骨(PC 36-10和PC 36-11)。這是亞洲古近紀已知的最好的一批松鼠化石標本,代表松鼠科Kherem屬的一新種:亞洲克熱姆鼠(Kherem asiatica sp。nov。)。其主要特徵是頰齒較大,較寬;下內尖的高度和下次尖的大小從m1到m3變化不大,彼此相近;下內脊較明顯;下內尖較明顯,與下後邊脊間有淺溝分開等。K。asiatica的形態特徵表明,Kherem屬應歸入北美的Cedromurinae亞科。新的發現表明,亞洲漸新世的松鼠已發生了分異,至少可分為3支。以Kherem為代表的一支,與北美早期的cedromurines有較近的系統關係。它可能是由類似於北美Oligospermophilus的種類演化並遷徙到亞洲來的。
山河狸
現生的山河狸只有一種(Aplodontia rufa),僅生活在北美西部沿海地區。但從始新世到中新世,山河狸卻遍布全北區。它們在北美較繁盛,分異大,但在亞洲發現得很少。在亞洲漸新世地層中雖曾報導過3屬4種山河狸化石,但材料都很少(prosciurus ordosicus和P。?shantun-gensis均只有一枚單個的牙)。山河狸化石在蒙古漸新世地層的新發現表明,亞洲漸新世的山河狸化石比已知的要豐富。該山河狸化石分別采自蒙古南戈壁省Shunkt地點的山地段和前杭愛省UlaanKhongil(=Tatal Gol)地點的塔塔爾段。它們代表原松鼠屬的3類:蒙古原松鼠(新種)(Prosci-urus mongoliensis sp。nov。)、蒙古原松鼠(相似種)(P。cf。P。mongoliensis)和小原松鼠(新種)(P。pisinnus sp。nov。)。蒙古原松鼠的主要特徵是下頰齒的下中附尖與下後附尖脊連,下中尖發育較弱,下外中脊弱或無;p4無下前邊尖和下前齒帶;ml具從下外脊伸向下次脊的小刺;m2和m3的下後脊Ⅱ較發育;下次脊在p4和m1後彎,與下後邊脊連,在m2和m3橫向,伸達下外脊等。
蒙古原松鼠(相似種)與蒙古原松鼠的區別是:頰齒尺寸較小,p4具明顯的下外中脊,p4和m1的下次脊彎曲度較緩,m1缺從下外脊伸向下次脊的小刺,m2~3的下後脊Ⅱ較短等。小原松鼠的主要特徵是:個體較小;齒冠較低;下臼齒的下中附尖孤立;下頰齒的下次脊橫向、較低,與下外脊連,下中尖和下外中脊均較明顯,下次尖明顯向前頰側延伸,具明顯的齒帶等。蒙古漸新世嚙齒類化石的新發現表明,亞洲漸新世的松鼠和山河狸與北美的關係要比與歐洲的近。
海洋
海洋持續變冷,特別是在南極洲附近。
災難事件
有記錄的外星體撞擊事件:加拿大,努納武特,2,300萬年前,隕石坑直徑24千米。
超級火山爆發事件:美國,科羅拉多,LaGaritaCaldera超級火山,2,780萬年前,噴發量大於5,000km³,火山爆發指數9.2。
地質年代參照表
宙代紀世年代開始
百萬年前(GSSP)主要事件
顯生宙新生代新近紀全新世0.011430±0.00013人類繁榮
更新世1.806±0.005冰河時期,大量大型哺乳動物滅絕
人類進化到現代狀態
上新世5.332±0.005人類的人猿祖先出現
中新世23.03±0.05?
古近紀漸新世33.9±0.1大部份哺乳動物目崛起
始新世55.8±0.2?
古新世65.5±0.3?
中生代白堊紀??恐龍的繁榮和滅絕
白堊紀-第三紀滅絕事件,地球上45%生物滅絕
有胎盤的哺乳動物出現
侏羅紀?199.6±0.6?
三疊紀?251.0±0.7?
古生代二疊紀??二疊紀滅絕事件,地球上95%生物滅絕
盤古大陸形成
石炭紀?359.2±2.5?
泥盆紀?416.0±2.8?
志留紀?443.7±1.5?
奧陶紀?488.3±1.7?
寒武紀?542.0±1.0?
元古宙新元古代成冰紀?630+5/-30多細胞生物出現
拉伸紀?850?
中元古代狹帶紀?1000?
延展紀?1400?
蓋層紀?1600?
古元古代固結紀?1800?
造山紀?2050?
層侵紀?2300?
成鐵紀?2500?
太古宙新太古代??2800第一次冰河期
中太古代??3200?
古太古代??3600藍綠藻出現
始太古代??3800?
冥古宙早雨海代???地球上出現第一個生物---細菌
酒神代???古細菌出現
原生代???地球上出現海洋
隱生代???地球出現
地質時代百科
地質年代參照表
| 宙|宙 | 代 | 紀 | 世 | 年代開始 百萬年前(GSSP) | 主要事件 |
| 顯生宙 | 新生代 | 新近紀 | 全新世 | 0.011430 ± 0.00013 | 人類繁榮 |
| 更新世 | 1.806 ± 0.005 | 冰河時期,大量大型哺乳動物滅絕 人類進化到現代狀態 | |||
| 上新世 | 5.332 ± 0.005 | 人類的人猿祖先出現 | |||
| 中新世 | 23.03 ± 0.05 | ||||
| 古近紀 | 漸新世 | 33.9 ± 0.1 | 大部份哺乳動物目崛起 | ||
| 始新世 | 55.8 ± 0.2 | ||||
| 古新世 | 65.5 ± 0.3 | ||||
| 中生代 | 白堊紀 | 99.6 ± 0.9 | 恐龍的繁榮和滅絕 白堊紀-第三紀滅絕事件,地球上45%生物滅絕 有胎盤的哺乳動物出現 | ||
| 侏羅紀 | 199.6 ± 0.6 | 有袋類哺乳動物出現 鳥類出現 裸子植物繁榮 被子植物出現 | |||
| 三疊紀 | 251.0 ± 0.7 | 恐龍出現 卵生哺乳動物出現 | |||
| 古生代 | 二疊紀 | 299.0 ± 0.8 | 二疊紀滅絕事件,地球上95%生物滅絕 盤古大陸形成 | ||
| 石炭紀 | 359.2 ± 2.5 | 昆蟲繁榮 爬行動物出現 煤炭森林 裸子植物出現 | |||
| 泥盆紀 | 416.0 ± 2.8 | 魚類繁榮 兩棲動物出現 昆蟲出現 種子植物出現 石松和木賊出現 | |||
| 志留紀 | 443.7 ± 1.5 | 陸生的裸蕨植物出現 | |||
| 奧陶紀 | 488.3 ± 1.7 | 魚類出現;海生藻類繁盛 | |||
| 寒武紀 | 542.0 ± 1.0 | 寒武紀生命大爆炸 | |||
| 元古宙 | 新元古代 | 埃迪卡拉紀 | 630 +5/-30 | 多細胞生物出現 | |
| 成冰紀 | 850 | 發生雪球事件 | |||
| 拉伸紀 | 1000 | 羅迪尼亞古陸形成 | |||
| 中元古代 | 狹帶紀 | 1200 | |||
| 延展紀 | 1400 | ||||
| 蓋層紀 | 1600 | ||||
| 古元古代 | 固結紀 | 1800 | |||
| 造山紀 | 2050 | ||||
| 層侵紀 | 2300 | ||||
| 成鐵紀 | 2500 | ||||
| 太古宙 | 新太古代 | 2800 | 第一次冰河期 | ||
| 中太古代 | 3200 | ||||
| 古太古代 | 3600 | 藍綠藻出現 | |||
| 始太古代 | 3800 | ||||
| 冥古宙 | 早雨海代 | 地球上出現第一個生物---細菌 | |||
| 酒神代 | 古細菌出現 | ||||
| 原生代 | 地球上出現海洋 | ||||
| 隱生代 | 地球出現 | ||||
