
科學分類
界: 動物界 Animalia
 山河狸
山河狸門: 脊索動物門 Chordata
綱: 哺乳綱 Mammalia
目: 齧齒目 Rodentia
亞目: 松鼠形亞目 Sciuromorpha
物種簡介
齧齒目最原始的山河狸科現存唯一的1種,山河狸科動物大約在5000萬年前的始新世就已出現。山河狸因外形、習性與河狸有很多相似之處(如眼小、耳短、被毛短密而色暗,穴居水邊,修排水渠,以及咬食樹木枝杈等)而得名。
外形特徵
體型中等,體長30~46厘米;全身被覆暗灰色或赤褐色短絨毛;尾甚短,僅端部稍露出毛被外;體胖,重約1千克;頭短鈍,多須,額部凸圓;腿短粗,前後肢5趾,爪狹長;適於挖土,前肢拇趾具蹄狀爪,其他4趾都能握食物。在形態上仍保留著一些較原始的特徵,如頭骨上無眶後突,咀嚼肌中顳肌較強大等。
生活習性
生活于海拔2200米以下的森林和茂密灌叢下常年積水的窪地中。棲居地面有許多洞口和扇形土丘,洞口常用土封堵,地下有
 山河狸
山河狸生長繁殖
每年冬末或早春繁殖1次,妊娠期約1個月,一般產2~3仔,多達6仔。幼鼠10日後睜眼,於6月底出洞活動,2年後成熟。數量多時對林業有一定程度的危害,有時也偷食莊稼和危害河渠堤壩。
分布範圍
山河狸科Aplodontidae是現存最原始的齧齒目成員,擁有一些原始特徵。山河狸科史前有較多種類,而現存僅山河狸Aplodontia rofa一種,分布於北美洲西北部的濕潤的海岸針葉林中。山河狸體長30~40厘米,外形似小型的河狸,但是沒有河狸那樣扁平的尾巴,其尾長僅2.5厘米。山河狸食性廣泛,可以食用多種不同的植物,並有儲存食物的習慣。山河狸調節體溫和保持濕度的能力均較差,需要冬眠和夏蜇,平時則挖掘複雜的洞穴居住。
種群現象
松鼠科Sciuridae是一個非常成功的科,其種類繁多,分布廣泛,適應從半荒漠、高山到熱帶雨林的多種不同生活
 北美土撥鼠
北美土撥鼠 山河狸總科
山河狸總科鼠類動物,從5 300萬年前在地球上出世以來,到如今已有了29科430屬1814種生活在今天的地球上,約占當今世界哺乳動物屬種的40%。鼠類個體數量也是多得驚人,世界上每6個哺乳動物中就有一個是鼠。鼠類生活在除南極以外的世界每個角落,在陸上有跑的老鼠,跳的跳鼠,地下挖洞的鼢鼠;有生活在水裡的河狸,還有樹間飛的鼯鼠。鼠的形態、相貌也千差萬別,逗人喜愛的小松鼠,討厭的家耗子,頭上生角的上古魯鼠。但是不管差別多大,在生物學上統歸一類,叫齧齒目。它們共同的特點就是長了一對終生生長的大門牙,不停地在啃(齧)東西。這門牙不斷生長,如果任其生長,越來越長的門牙就會撐得連嘴巴都閉不上,這就是老鼠啃木箱的原因--並不為吃,只是為磨。有人把家鼠養在瓷缸內,飼以軟食,隔些天后,鼠嘴被門牙頂撐,無法合攏進食而致死。其實,鼠類在吃東西時,是用門牙把食物咬斷後,送到後面用臼齒咀嚼,這時門牙是張開的,這一特點是鼠類所特有的。為了咀嚼方便,鼠的犬齒沒了,頰齒數目也大大減少。
瀕危狀況
中等,尚無滅絕危險
保護措施
尚無對該物種的保護
物種價值
鼠類對人類生活甚為重要,它偷糧,傳播鼠疫,破壞農田和草原。但它的毛皮可用,而且它繁殖快、數量多,人們常把它用作實驗動物,稱之為“哺乳動物中的果蠅”。
物種進化演變史
齧齒目Rodentia是哺乳動物中最大最成功的一類,擁有哺乳動物中2/5的種類,分布幾乎遍及南極和少數海島以外的世界各地。對於種類繁多的嚙齒類成員之間的親緣關係爭議比較大,主要是根據顱骨上咬肌的結構和附著情況、
 松鼠
松鼠五花八門的齧齒目在生物學上根據牙齒、咬肌結構大體分成4大類:始齧類(如今僅在北美殘留一種--山河狸)、松鼠類(包括松鼠和河狸)、鼠類(家鼠、倉鼠和跳鼠)和豚鼠類(豚鼠和水豚)。近年來分子生物學的研究認為,南美豚鼠與其他嚙齒類的相似性低於它與靈長類或其他哺乳動物的相似性,因此認為南美豚鼠不是嚙齒類,從而對傳統的嚙齒類概念提出挑戰。
始齧類繁盛在古近紀,到漸新世地球驟冷時基本上滅絕了。大間斷後一些現生的科出現了,如河狸、松鼠、跳鼠和南美豚鼠等。鼠科動物是最龐大的,它包括倉鼠、
 倉鼠
倉鼠由於嚙齒類種群的數量多,進化快,是確定地層時代最為有效的指示化石。以一種叫模鼠的田鼠類化石為例,在不到300萬年的時間裡,牙齒從有根到無根,齒面的紋飾也逐漸變化。如果在地層里找到哪種模鼠,就可以推斷該地層的時代了。
嚙齒類的起源,一直是多年未解的問題。過去總以北美早始新世的副鼠為樣板,推測嚙齒類起源於古新世的靈長類。近年在我國發現的新材料證實,嚙齒類應當起源於中亞:安徽潛山古新世地層中的東方曉鼠是嚙齒類的祖先類型,內蒙古的磨楔齒鼠則是最原始的嚙齒類。
從20世紀20年代起,動物學界一直認為鼠和兔沒有系統關係,兔類可能起源於偶蹄類、古老的踝節類或狃獸類。直到70年代在安徽潛山與曉鼠一起發現了兔形類的祖先--模鼠兔,才算重新確認了鼠和兔的親緣關係,兩者都歸入同一大類--齧形超目。兔和鼠都長有一對無根的大門牙,只是兔類在大門牙後面還有一對小門牙,所以在分類上把兔類也叫雙門齒目,而嚙齒類則為單門齒目。
飼養方法及注意事項
該物種尚未被人類馴化。
相關詞條
參考文獻
重大報導
蒙古新發現的漸新世山河狸和松鼠類化石 蒙古中部地區的三達河組(Hsanda Gol Formation)是亞洲漸新世陸相地層的經典地層之一.三達河組的沉積岩層現被分成兩個岩性段:上部早漸新世晚期,晚漸新世山地段(Shand Member)和下部早漸新世早期塔塔爾段(Tatal Member);兩岩段間有時夾有Khar Khoroo玄武岩層(Bryant and McKenna,1995;Meng and McKenna,1998).該玄武岩層的時代用40Ar/39Ar法被確定為距今31.5 Ma(Hock et al.,1999,他們稱其為Basalt Ⅰ).三達河組中產有豐富的哺乳動物化石,特別是嚙齒類化石(Matthew and Granger,1923;Kowalski,1974;Russelland Zhai,1987;Hock et al.,1999).近年來,本文的後一作者在三達河組中又採集到一批嚙齒類化石,其中包括松鼠和山河狸2科的化石.
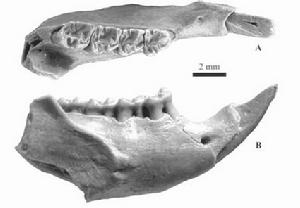 發現的山河狸頭骨化石
發現的山河狸頭骨化石山河狸化石分別采自蒙古南戈壁省Shunkt 地點的山地段和前杭愛省UlaanKhongil 地點的塔塔爾段。它們代表原松鼠屬的3 類:蒙古原松鼠(新種) 和小原松鼠(新種)。蒙古原松鼠的主要特徵是下頰齒的下中附尖與下後附尖脊連,下中尖發育較弱,下外中脊弱或無;p4 無下前邊尖和下前齒帶;m1 具從下外脊伸向下次脊的小刺;m2 和m3 的下後脊II 較發育;下次脊在p4 和m1 後彎,與下後邊脊連,在m2 和m3 橫向,伸達下外脊等。蒙古原松鼠(相似種) 與蒙古原松鼠的區別是:頰齒尺寸較小,p4 具明顯的下外中脊,p4 和m1 的下次脊彎曲度較緩,m1 缺從下外脊伸向下次脊的小刺,m2~3 的下後脊II較短等。小原松鼠的主要特徵是:個體較小;齒冠較低;下臼齒的下中附尖孤立;下頰齒的下次脊橫向、較低,與下外脊連,下中尖和下外中脊均較明顯,下次尖明顯向前頰側延伸,具明顯的齒帶等。蒙古漸新世嚙齒類化石的新發現表明,亞洲漸新世的松鼠和山河狸與北美的關係要比與歐洲的近。
相關連線
http://www.csdyzx.com/swtd/dongwu/jzdwm/frdwm23.htm
http://baike.baidu.com/view/45498.htm
http://zh.wikipedia.org/w/index.php?title=%E6%9D%BE%E9%BC%A0%E5%BD%A2%E4%BA%9E%E7%9B%AE&variant=zh-cn
動物的世界2
| 動物是生物界中的一大類。一般不能將無機物合成有機物,只能以有機物(植物、動物或微生物)為食料,因此具有與植物不同的形態結構和生理功能,以進行攝食、消化、吸收、呼吸、循環、排泄、感覺、運動和繁殖等生命活動。我們一起來研究研究這些動物吧! |
