
定義
分泌蛋白(secretedprotein)
定義:是指在細胞內合成後,分泌到細胞外起作用的的蛋白質。例如:唾液澱粉酶,胃蛋白酶,消化酶,抗體和一部分激素。註:例如呼吸酶就不屬於分泌蛋白。
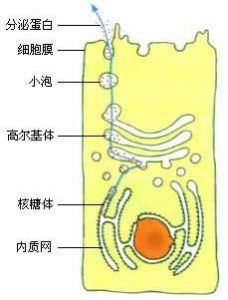
在核糖體上合成的分泌蛋白,要經過內質網和高爾基體,而不是直接運輸到細胞膜。
 分泌蛋白
分泌蛋白進一步的研究表明,在核糖體上翻譯出的蛋白質,進入內質網腔後,還要經過一些加工,如摺疊、組裝、加上一些糖基團等,才能成為比較成熟的蛋白質。然後,由內質網腔膨大、出芽形成具膜的小泡,包裹著蛋白質轉移到高爾基體,把蛋白質輸送到高爾基體腔內,做進一步的加工。接著,高爾基體邊緣突起形成小泡,把蛋白質包裹在小泡里,運輸到細胞膜,小泡與細胞膜融合,把蛋白質釋放到細胞外。
蛋白質的引導
蛋白質的運輸儘管比較複雜,但是生物系統中的蛋白質的運輸可以用一個比較簡單的模式來解釋。每個需要運輸的多肽都很有一段胺基酸序列,稱為信號肽序列,引導多肽到不同的轉運系統。
 分泌蛋白的靶向輸送
分泌蛋白的靶向輸送信號肽及其作用機制
定義
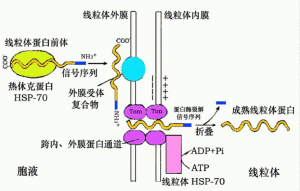
70年代初期,許多研究發現,在編碼分泌蛋白的基因中,許多基因的5'端都有一段DNA編碼的15~35個胺基酸的疏水性肽片段,這一位於蛋白質N——末端的肽段在成熟的分泌蛋白中並不存在,其功能在於引導隨後產生的蛋白質多肽鏈穿過內質網膜進入腔內。這一段疏水性短肽在蛋白質的內質網——高爾基體——質膜分泌途徑中具有重要作用,並被稱之為信號肽。
學術觀點
1975年,布洛貝爾提出了信號肽假說。根據這一假說,在細胞質中,編碼分泌蛋白的信使核糖核酸(mRNA)與游離的核糖體大小亞基結合而形成翻譯複合體。從起始密碼子開始,首先翻譯產生信號肽,當轉譯進行到大約50~70個胺基酸之後,信號肽開始從核糖體的大亞基上露出,露出的信號肽立即被細胞質中的信號肽識別體(SRP)識別並與之相結合。此時,轉譯暫時停止,SRP牽引這條帶核糖體的mRNA到達粗面內質網的表面,並與粗面內質網表面上的信號肽識別體受體(或稱停泊蛋白)作用,這時,暫時被抑制的轉譯過程恢復進行,同時,內質網膜上某種特定的核糖體受體蛋白聚集,使膜雙脂層產生孔道,帶mRNA的核糖體與其受體蛋白結合,轉譯出的肽鏈便通過孔道進入內質網腔內。
原理
信號肽在穿越膜後即被內質網腔內的信號肽酶水解切除。當核糖體與其受體蛋白結合後,SRP與停泊蛋白便解離,各自進入新的識別、結合循環。當轉譯進行到mRNA的終止密碼子時,蛋白質的合成結束,核糖體的大小亞基解聚,大亞基與核糖體受體的相互作用消失,核糖體受體解聚,內質網膜上的蛋白孔道消失,內質網恢復成完整的脂雙層結構。進入內質網腔內的多肽鏈在信號肽被水解切除後即進行摺疊及其他一系列修飾過程,最終形成成熟的分泌蛋白。
分泌蛋白在內質網(ER)內合成
概述
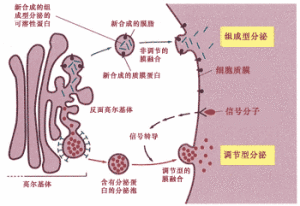
在真核細胞中,內質網是最大的膜狀結構的細胞器,其表面積可以是質膜面積的幾倍。大部分的內質網與核糖體相結合形成糙面內質網,在糙面內質網上的核糖體是膜蛋白和分泌蛋白合成的地方,也是蛋白質分泌途徑的起點。多肽經移位後,在內質網的小腔中被修飾,通過短時間的加工後,分泌蛋白形成被膜包裹的小泡,轉運到高爾基體,然後再轉運到細胞表面或溶酶體。
 分泌蛋白的形成
分泌蛋白的形成1、蛋白質的修飾與加工
這些修飾包括糖基化、羥基化、醯基化、二硫鍵形成等,其中最主要的是糖基化,幾乎所有內質網上合成的蛋白質最終被糖基化。糖基化的作用是:①使蛋白質能夠抵抗消化酶的作用;②賦予蛋白質傳導信號的功能;③某些蛋白只有在糖基化之後才能正確摺疊。糖基化有兩種類型:(1)糖蛋白是由寡糖連線在Asp的氨基的形成的,連線的鏈叫N-糖苷鍵。(2)寡糖連線在Ser、Thr或羥基-lys的羥基上(O-糖苷鍵)。N-糖苷鍵是在ER開始,而在高爾基體中進一步完成;O-糖苷鍵的形成僅發生在高爾基體中。N-糖基化可分為3步,各種N-連線的寡糖都是在ER中開始加上的,途徑也相同。一個寡糖含有2個N-乙醯-糖胺,9個苷露糖和3個葡萄糖,在一種特異的脂一多萜醇上形成,多萜醇磷酸酯即是攜帶糖的載體。多萜醇是一種高度疏水的酯,位於ER膜中,其活性基團面向著ER腔,寡糖是由單個的糖連線而構成,它通過焦磷酸和多萜醇連線。寡糖作為一個單位在與膜結合的糖基轉移酶的作用下從多萜醇上轉移到靶蛋白上。酶的活性位點也是露在ER腔中。受體基團是位於Asn-X-Ser或Asn-X-Thr(X是除Pro以外的任何胺基酸)中的Asn,當新生肽進入ER時,它一旦被識別後立即作為靶順序暴露在腔中。有些寡糖的修整是在ER中進行,修整後再進入高爾基體。寡糖結構完成是分為兩類,一類在轉運到ER時,另一類是在越膜轉運到高爾基體。究竟屬於何種類型這要取決於苷露糖。苷露糖只是在ER中加上,隨後可能還要被修整。在ER中被切去的單糖的數目是不同的。ER中的苷露糖苷酶很快地附著到第一個苷露糖上,附著下三個較慢。高甘露糖寡聚糖含有的殘基是在ER中加上的。寡糖加上後幾乎立即又從蛋白上被切掉。3個葡萄糖殘基被ER中的葡萄糖苷酶切除掉,ER中的甘露糖苷酶再從蛋白上切下2-4個甘露糖,在ER中產生3寡糖的最終結構。
2、新生肽鏈的摺疊、組裝,運輸
在ER腔中折選和修飾是有關的,糖的連線對於正確的摺疊是十分必要的。蛋白二硫異構酶可以改變二硫鍵,影響到摺疊,它和特殊的ER蛋白的結合是必須的,此酶的某些活性或全部的活性可能是酶作為ER中的一種複合體的形式來實現的。即在越膜位點和蛋白結合才能發揮它的功能。多肽經過內質網的加工、修飾、摺疊後被膜包裹形成小泡轉運到高爾基體在高爾基體進行進一步的加工。
在高爾基體的進一步加工
高爾基體的主要有兩方面的功能:一是對糖蛋白上的寡糖核作進一步的修飾與調整,二是將各種多肽進行分類並送往溶酶體、分泌粒和質膜的功能目的地。但蛋白質應送往哪裡是由蛋白質本身的空間結構決定的。
高爾基體是由許多層袋狀的膜結構組成的。糖蛋白的進一步糖基化修飾就是在這種膜結構中完成的,如複合寡聚糖就是在高爾基體中進一步修整和加上糖的殘基。第一步是通過高爾基體的甘露糖苷酶Ⅰ修整甘露糖殘基。然後單個的糖基由N-乙醯-葡萄糖胺轉移加上,按著由高爾基體苷露糖苷酶Ⅱ繼續切除苷露糖殘基。
在高爾基體的修飾中都會產生內部核心,它是由NAc-Glc·NAc-GLc.Man3構成,最後要被剝去。末端區域加在內部核心下。末端區域的殘基包括NAc-GLc,Gal和唾液酸(N-乙醯-神經氨[糖]酸。此加工的路徑和糖基化是高度有序的,而且有兩種類型的反應。一種糖殘基的加入對於另一種糖基的剪下可能是必要的。如在最終剪除甘露糖之前要加上NAc-Glc。
現在還不知道加工過程中各種蛋白的降解,加工的模式及糖基化的信號是什麼。據推測此信號在肽鏈的結構中,而不可能在寡糖中,因為N-糖苷鍵開始形成都是加上相同的寡糖。
經過高爾基體的進一步加工和分裝,成熟的蛋白質通過小泡運到細胞表面或者是溶酶體。伴隨各種具膜小泡的運輸過程,細胞內形成了複雜的膜流,高爾基體在在膜流的調控中起樞紐的作用。
沒有活性的前體蛋白,進行一個系列的翻譯後加工後才能成為具有功能的成熟蛋白,各胞器之間有的明顯分工但是個細胞器在分泌蛋白的形成和分泌過程中的又是相互聯繫密切相關的。
