
簡介
內質網(ER,Endoplasmic Reticulum),細胞質內由膜組成的一系列片狀的囊腔和管狀的腔,彼此相通形成一個隔離於細胞基質的管道系統,為細胞中的重要細胞器。實際上是一個連續的膜囊和膜管網,可分為糙(粗)面內質網(RER, Rough Endoplasmic Reticulum)和光(滑)面內質網(SER, Smooth Endoplasmic Reticulum)兩大部分。
糙面內質網(ergastoplasm),也稱為粗面內質網或顆粒型內質網
是指腺細胞和生長中的卵母細胞等的固定材料,在光學顯微鏡下呈線狀、層狀,可被鹼性染料充分染色的為C.Garnier(1897)稱之為細胞質的部分。後來證明其嗜鹼性是由於RNA的存在所決定的。尤其由於電子顯微鏡的發展,顯示該部分富有粗面內質網,並了解到RNA的存在是由內質網上的核糖體決定的。從而肯定了粗面內質網是細胞內蛋白質的生物合成場所這一假想。
蛋白質合成
(一)蛋白質合成蛋白質都是在核糖體上合成的,並且起始於細胞質基質,但是有些蛋白質在合成開始不久後便轉在內質網上合成,這些蛋白質主要有:
①向細胞外分泌的蛋白、如抗體、激素;
②跨膜蛋白,並且決定膜蛋白在膜中的排列方式;
③需要與其它細胞組合嚴格分開的酶,如溶酶體的各種水解酶;
④需要進行修飾的蛋白,如糖蛋白。
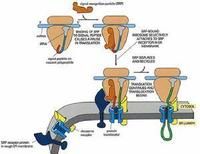
C. Milstein(1972)發現從骨髓瘤細胞提取的免疫球蛋白分子N端要比分泌到細胞外的N端多出一段。G. Blobel和D. Sabatini等根據進一步的實驗,提出了信號假說(Signal hypothesis),認為蛋白質上的信號肽,指導蛋白質轉至內質網上合成。Blobel因此項發現獲1999年諾貝爾生理醫學獎。
蛋白質轉入內質網合成至少涉及5種成分:
①信號肽(signal peptide),是引導新合成肽鏈轉移到內質網上的一段多肽,位於新合成肽鏈的N端,一般16~30個胺基酸殘基,含有6-15個帶正電荷的非極性胺基酸,由於信號肽又是引導肽鏈進入內質網腔的一段序列,又稱開始轉移序列(start transfer sequence)。
②信號識別顆粒(signal recognition particle,SRP),由6種結構不同的多肽組成,結合一個7S RNA,分子量325KD,屬於一種核糖核蛋白(ribonucleoprotein)。SRP與信號序列結合,導致蛋白質合成暫停。
③ SRP受體(SPR receptor),是膜的整合蛋白,為異二聚體蛋白,存在於內質網上,可與SRP特異結合。
④停止轉移序列(stop transfer sequence),肽鏈上的一段特殊序列,與內質網膜的親合力很高,能阻止肽鏈繼續進入內質網腔,使其成為跨膜蛋白質。
⑤轉位因子(translocator),由3-4個Sec61蛋白複合體構成的一個類似炸面圈的結構,每個Sec61蛋白由三條肽鏈組成。
蛋白質轉入內質網合成的過程:
信號肽與SRP結合→肽鏈延伸終止→SRP與受體結合→SRP脫離信號肽→肽鏈在內質網上繼續合成,同時信號肽引導新生肽鏈進入內質網腔→信號肽切除→肽鏈延伸至終止→翻譯體系解散。這種肽鏈邊合成邊向內質網腔轉移的方式,稱為co-translation。
一些信號肽序列的蛋白質及信號序列
Preproalbumin
Met-Lys-Trp-Val-Thr-Phe-Leu-Leu-Leu-Leu-Phe-Ile-Ser- Gly-Ser-Ala-Phe-Ser↓Arg...
Pre-IgG light chain
Met-Asp-Met-Arg-Ala-Pro-Ala-Gln-Ile-Phe-Gly-Phe-Leu- Leu-Leu-Leu-Phe-Pro-Gly- Thr-Arg-Cys↓Asp...
Prelysozyme
Met-Arg-Ser-Leu-Leu-Ile-Leu-Val-Leu-Cys-Phe-Leu- Pro-Leu-Ala-Ala-Leu-Gly↓Lys...
包括糖基化、羥基化、醯基化、二硫鍵形成等,其中最主要的是糖基化,幾乎所有內質網上合成的蛋白質最終被糖基化。糖基化的作用是: ①使蛋白質能夠抵抗消化酶的作用;②賦予蛋白質傳導信號的功能;③某些蛋白只有在糖基化之後才能正確摺疊。
糖基一般連線在4種胺基酸上,分為2種:
O-連線的糖基化(O-linked glycosylation):與Ser、Thr和Hyp的OH連線,連線的糖為半乳糖或N-乙醯半乳糖胺,在高爾基體上進行O-連線的糖基化。
N-連線的糖基化(N-linked glycosylation):與天冬醯胺殘基的NH2連線,糖為N-乙醯葡糖胺。
內質網上進行的為N-連線的糖基化。糖的供體為核苷糖(nucleotide sugar),如CMP-唾液酸、GDP-甘露糖、UDP-N-乙醯葡糖胺等。糖分子首先被糖基轉移酶轉移到膜上的磷酸長醇(dolichol phosphate)分子上,裝配成寡糖鏈。再被寡糖轉移酶轉到新合成肽鏈特定序列(Asn-X-Ser或Asn-X-Thr)的天冬醯胺殘基上。
(三)新生肽鏈的摺疊、組裝和運輸
 蛋白質轉移到內質網上合成的過程
蛋白質轉移到內質網上合成的過程COP II介導由內質網輸出的膜泡運輸,這種膜泡由內質網的排出位點(exit sites)以出芽的方式排出,內質網的排出位點沒有結合核糖體,隨機分布在內質網上。不同的蛋白質在內質網腔中停留的時間不同,主要取決於蛋白質完成正確摺疊和組裝的時間,這一過程是在屬於hsp70家族的ATP酶的作用下完成的,需要消耗能量。有些無法完成正確摺疊的蛋白質被輸出內質網,轉入溶酶體中降解掉,大約90%的新合成的T細胞受體亞單位和乙醯膽鹼受體都被降解掉,而從未到達靶細胞膜。
病變
1、粗面內質網擴張、囊泡化正常粗面內質網系由膜形成扁池,腔很窄,在細胞水腫時,液體入腔內使之擴張,並形成小泡,在肝炎時肝細胞氣球樣變中也可見到。
2、粗面內質網脫粒粗面內質網扁池膜旁有核糖體依附,在粗面內質網腫脹同時膜表面核糖體脫落。也有的粗面內質網膜表面核糖體脫落,但扁池不擴張。
3、粗面內質網板層在內分泌細胞及胚胎肝細胞內出現短而緊密排列粗面內質網,在內分泌腫瘤及肝癌中也可見到。
4、同心圓膜性小體有三種:第一種粗面內質網呈同心圓板層排列;第二種是滑面膜雙層呈同心圓排列;第三種是在滑面間夾有糖原。
5、粗面內質網池內隔離粗面內質網扁池擴張,帶有核糖體的膜突入擴張的池內,切面如像島狀膜性小管小泡游離在池內。
6、粗面內質網對合池平行兩片粗面內質網緊密靠攏,內側面核糖體消失。也可有三片或多片,稱三合池或多合池。
