
簡介
 順式作用元件
順式作用元件順式作用元件在分子遺傳學領域,相對同一染色體或DNA分子而言為“順式”(cis);對不同染色體或DNA分子而言為“反式”(trans)。
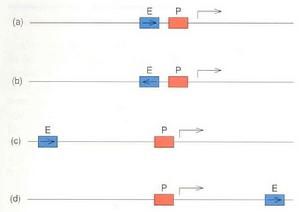
順式作用元件是轉錄調節因子的結合位點,包括啟動子、增強子和沉默子。真核基因啟動子是原核啟動序列的同義語。真核啟動子是指RNA聚合酶及轉錄起始點周圍的一組轉錄控制組件,每個啟動子包括至少一個轉錄起始點以及一個以上的功能組件,轉錄調節因子即通過這些機能組件對轉錄起始發揮作用。在這些調節組件中最具典型意義的就是TATA盒子,它的共有序列是TATAAA。TATA盒子通常位於轉錄起始點上游-25至-30區域,控制轉錄的準確性和頻率。TATA盒子是基本轉錄因子TFⅡD結合位點;TFⅡD則是RNA聚合酶結合DNA必不可少的。除TATA盒子外,GC盒子(GGGCGG)和CAAT盒子(GCCAAT)也是很多基因中常見的,它們位於起始點上游-30至-110bp區域。所謂增強子就是遠離轉錄起始點、決定組織特異性表達、增強啟動子轉錄活性的特異DNA序列,其發揮作用的方式與方向、距離無關。增強子與啟動子非常相似:都是由若干組件組成,有些組件既可在增強子、又可在啟動子出現。從功能方面講,沒有增強子存在,啟動子通常不能表現活性;沒有啟動子,增強子也無法發揮作用。增強子和啟動子有時分隔很遠,有時連續或交錯覆蓋。
某些基因有負性調節元件棗抑制子(沉默子)存在。有些DNA序列既可作為正性、又可作為負性調節元件發揮順式調節作用,這取決於不同類型細胞中DNA結合因子的性質。
結構組成
順式作用元件是同一DNA分子中具有轉錄調節功能的特異DNA序列。按功能特性,真核基因順式作用元件分為啟動子、增強子及沉默子。
啟動子
原核操縱子中啟動序列的同義語。真核基因啟動子是RNA聚合酶結合位點周圍的一組轉錄控制組件,每一組件含7~20bp的DNA序列。啟動子包括至少一個轉錄起始點以及一個以上的機能組件。在這些機能組件中最具典型意義的就是TATA盒,它的共有序列是TATAAAA。TATA盒通常位於轉錄起始點上游-25~-30bp,控制轉錄起始的準確性及頻率。TATA盒是基本轉錄因子TFIID結合位點。除TATA盒外,GC盒(GGGCGG)和CAAT盒(GCCAAT)也是很多基因常見的,它們通常位於轉錄起始點上游-30~-110bp區域。此外,還發現很多其它類型的機能組件。由TATA盒及轉錄起始點即可構成最簡單的啟動子。
增強子
就是遠離轉錄起始點(1~30kb)、決定基因的時間、空間特異性表達、增強啟動子轉錄活性的DNA序列,其發揮作用的方式通常與方向、距離無關。增強子也是由若干機能組件
 順式作用元件分布
順式作用元件分布增強子增強子的特點:
(1)增強子可提高同一條DNA鏈上基因轉錄效率,可以遠距離作用,通常距離l~4kb,個別情況下離開所調控的基因30kb仍能發揮作用,而且在基因的上游或下游都能起作用。
(2)增強子作用與其序列的正反方向無關,將增強子方向倒置依然能起作用。而將啟動子倒置就不能起作用,可見增強子與啟動子是很不相同的。
(3)增強子要有啟動子才能發揮作用,沒有啟動子存在,增強子不能表現其活性。但增強子對啟動子沒有嚴格的專一性,同一增強子可以影響不同類型啟動子的轉錄。
(4)增強子必須與特定的蛋白質因子結合後才能發揮增強轉錄的作用。增強子一般具有組織或細胞特異性,許多增強子只在某些細胞或組織中表現活性,是由這些細胞或組織中具有的特異性蛋白質因子所決定的。例如,人類胰島素基因5’端上游約250個核苷酸處有一組織特異性增強子。在胰島素p細胞中有一種特異性蛋白因子,可以作用於這個區域,以增強胰島素基因的轉錄。在其他組織細胞中沒有這種蛋白因子,所以也就沒有此作用。這就是為什麼胰島素基因只有在胰島素p細胞中才能很好表達的重要原因。
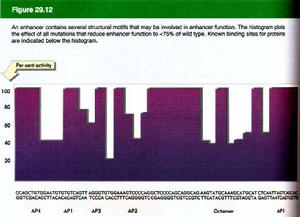
(5)大多為重複序列,一般長約50bp,適合與某些蛋白因子結合。其內部常含有一個核心序列,即(G)TGGA/TA/TA/T(G),是產生增強效應時所必需的。
(6)增強效應十分明顯,一般能使基因轉錄頻率增加10—200倍。經人巨大細胞病毒增強子增強後的珠蛋白基因表達頻率比該基因正常轉錄高600~1000倍。
(7)許多增強子還受外部信號的調控,如金屬硫蛋白的基因啟動區上游所帶的增強子,就可以對環境中的鋅、鎘濃度做出反應。
(8)增強子的功能是可以累加的。SV40增強子序列可以被分為兩半,每一半序列本身作為增強子功能很弱,但合在一起,即使其中間插入一些別的序列,仍然是一個有效的增強子。因此,要使一個增強子失活必須在多個位點上造成突變。對SV40增強子而言,沒有任何單個的突變可以使其活力降低10倍。
增強子的作用原理:一種觀點認為,增強子為轉錄因子提供進入啟動子區的位點。另一種認為,增強子能改變染色質的構象。因為增強子區域容易發生從B—DNA到Z—DNA的構象變化。
沉默子
某些基因含有的一種負性調節元件,當其結合特異蛋白因子時,對基因轉錄起阻遏作用。某些基因有負性調節元件棗抑制子(沉默子)存在。有些DNA序列既可作為正性、又可作為負性調節元件發揮順式調節作用,這取決於不同類型細胞中DNA結合因子的性質。元件作用
 順式作用元件
順式作用元件多細胞有機體在生長、分化和發育過程中需要整合不同組織的、發育的、環境的信號調節基因表達,轉錄起始的調節是其中的重要一環。順式作用(cisacting)元件是同一DNA分子中具有轉錄調節功能的特異DNA序列,即具有特殊功能的轉錄因子DNA結合位點和其他調控基序(motif)。
與反式作用因子關係
 順式作用元件實驗
順式作用元件實驗反式作用因子:指和順式作用元件結合的可擴散性蛋白,包括基礎因子,上游因子,誘導因子。
真核生物的轉錄調控是調控的最重要的途經,大多是通過順式作用元件和反式作用因子複雜的相互作用而實現的。順式作用元件(cis-actingelement)存在於基因旁側序列中能影響基因表達的序列,它們的作用是參與基因表達的調控,本身不編碼任何蛋白質,僅僅提供一個作用位點,要與反式作用因子相互作用而起作用。啟動子存在著很多順式元件,可分為以下四種類型:①核心啟動子成分,如TATA框;②上游啟動子成分,如CAAT框,GC框,ATF結合位點;③遠上游元件:如增強子,沉默子等;④特殊啟動子成分:如淋巴細胞中的Oct(octamer)和κB。這些元件都為不同的轉錄因子及蛋白質識別和結合來調節轉錄。
反式作用因子(trans-actingfactor)通過以下不同的途經發揮調控作用:蛋白質和DNA相互作用;蛋白質和配基結合;蛋白質之間的相互作用以及蛋白質的修飾。參與基因表達調控的因子,它們與特異的靶基因的順式元件結合起作用。編碼反式作用因子的基因與被反式作用因子調控的靶序列(基因)不在同一染色體上。反式作用因子有兩個重要的功能結構域:DNA結合結構域和轉錄活化結構域,它們是其發揮轉錄調控功能的必需結構。反式作用因子可被誘導合成,其活性也受多種因素的調節。同一類序列特異性的反式作用因子由多基因家族所編碼。
與轉錄水平調節的反式作用因子通常可分為四大類(RobertG.Roeder,2004):①RNA聚合酶;②和RNA聚合酶相聯繫的普遍性轉錄因子(generaltranscriptionfactor,GTFs)它們結合在靶基因的啟動子上,形成前起始複合物(pre-initiationcomplex,PIC),啟動基因的轉錄;③特異性轉錄因子(gene-specific(DNA-binding)regulatoryfactors)(如激活因子和抑制因子),一類與靶基因啟動子和增強子(或沉默子)特異結合的轉錄因子,具有細胞及基因特異性,可以增強或抑制靶基因的轉錄;④種類多樣的協調因子(coregulatoryfactors),要么改變局部染色質的構像(如組蛋白醯基轉移酶和甲基轉移酶),對基因轉錄的起始具有推動作用,要么直接在轉錄因子和前起始複合物之間發揮橋樑作用(如中介因子(Mediator)),推動前起始複合物(PIC)形成和發揮作用。包括:
1、螺鏇轉角螺鏇(Helix-turn-helix):有3個螺鏇,螺鏇3是識別和DNA結合,一般結合於大溝;螺鏇1和2和其它蛋白質結合。
2、鋅指結構:鋅指(zincfimger)這個名稱來源於它的結構,它由一小組保守的胺基酸和鋅離子結合,在蛋白質中形成了相對獨立的功能域,像一根根手指伸向DNA的大溝。兩種類型的DNA結合蛋白具有這種結構,一類是鋅指蛋白,另一類是甾類受體。
3、亮氨酸拉鏈(Leucineziipper)兩個蛋白之間相互作用共同建立一個轉錄複合體,在一系列轉錄因子中發現的一種模體涉及兩個相同和不同的部分構成。亮氨酸拉鏈是一種富含亮氨酸的蛋白鏈形成的二聚體模體。它本身形成二聚體同時還可以識到特殊的DNA序列。一個親水的α-螺鏇在其表面的一側有疏水基因(包括亮氨酸),另一側表面帶有電荷。
4、螺鏇一環一螺鏇(HLH)
5、同源異形結構域(Homeodomains,HD):同源異形盒(hoomeobox)是一種編碼60aa的序列,長180bp,幾乎存在於所有真核生物中。
