
敘述
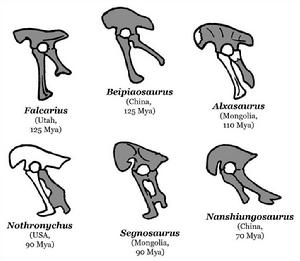 各種鐮刀龍超科的骨盆,其中鑄鐮龍可能不屬於鐮刀龍超科
各種鐮刀龍超科的骨盆,其中鑄鐮龍可能不屬於鐮刀龍超科鐮刀龍類擁有非常獨特的特徵,常會造成混淆。它們的長頸部、寬廣腹部、以及有四個腳趾的腳部,類似原蜥腳下目恐龍。它們的臀部骨頭往後,部分骨頭癒合者,這些特徵起初讓古生物學家以為是鳥臀目恐龍。
鐮刀龍類的最獨特特徵是手上的巨大指爪,鐮刀龍的指爪甚至可達3呎長。鐮刀龍類的不尋常前肢移動範圍,可讓它們的手觸及其他獸腳類恐龍不能摸到的地方,加上它們的牙齒特徵,許多古生物學家認為它們主要是草食性動物。鐮刀龍類可使用長而大幅彎曲的指爪來抓取並切碎樹枝,類似史前的大地懶。
北票龍的化石上具有皮膚痕跡,顯示鐮刀龍類覆蓋者原始的絨羽,類似美頜龍科的中華龍鳥;除此之外,北票龍還有第二型態羽毛,較長、較寬。鐮刀龍類的體型有很大的差距,小型的北票龍身長2.2米,而巨大的鐮刀龍身長接近10到12米,體重估計有6.2公噸,是最巨大的獸腳亞目恐龍之一。
發現歷史
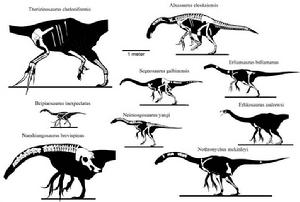 數種鐮刀龍超科已發現的部分
數種鐮刀龍超科已發現的部分因為早期發現的鐮刀龍類化石非常不完整,它們的奇異生理特徵讓某些科學家,例如葛瑞格利·保羅(Gregory S. Paul),認為慢龍類是存活到很晚期的原始恐龍,有時被認為是蜥臀目與鳥臀目的中間物種。因為慢龍類被認為與原蜥腳類有親緣關係,所以早期的描述(包括保羅的想像圖)將它們描繪成半四足動物;然而,對鐮刀龍類的類似鳥類腕部而言,這種行走方式是相當不可能的。除此之外,葛瑞格利·保羅曾將慢龍類歸類為“Phytodinosauria”(意為“植物恐龍”)的一個亞目;“Phytodinosauria”是古生物學家羅伯特· 巴克(Robert Bakker)在1986年建立,以包含鳥臀目、原蜥腳下目、蜥腳下目在內的草食性恐龍,共同特徵是牙齒鈍、呈匙狀,適合咬碎植物。
直到1990年代中期發現了阿拉善龍的化石,這些化石顯示出典型的獸腳類特徵,而鐮刀龍被重新分類於慢龍下目,因此才發現它們是群肉食性獸腳類恐龍的草食性後代。近年因為發現了原始的鐮刀龍類物種,例如1999年的北票龍與2005年的鑄鐮龍,而更清楚衍化的鐮刀龍科與其他獸腳類之間的關係。研究鑄鐮龍的科學家注意到,鑄鐮龍似乎代表者肉食性手盜龍類與草食性鐮刀龍類之間的遺失環節。雖然鐮刀龍超科被重新分類於獸腳亞目,但它們類似蜥腳下目的頭顱骨,以及牙齒與頜部的形狀讓它們極可能是草食性動物。
分類學
分類
 北票龍的重建圖
北票龍的重建圖瑞欽·巴思缽(Rinchen Barsbold)與Perle在1980年建立了慢龍下目(Segnosauria),屬於獸腳亞目。在1992年,董枝明進一步將慢龍類改列為獨自的慢龍目(Segnosaurischia)。但自從慢龍類被發現屬於獸腳亞目之後,這名稱遭到廢棄,卻也沒有過系統發生學上的定義;直到2004年,Clark等人認為慢龍下目是鐮刀龍超科的同義詞。
鐮刀龍超科是由葉甫根尼·馬列夫(Evgeny Maleev)在1954年所建立,當時僅包含擁有巨大指爪的奇特獸腳類恐龍鐮刀龍。之後當鐮刀龍被發現是種先進的慢龍類,鐮刀龍超科開始有了系統發生學上的定義,並逐漸取代了慢龍下目,主要原因是慢龍下目這名稱會讓人誤認為是原蜥腳下目的近親。
以下來自於Lindsay E. Zanno的2010年鐮刀龍類重新研究:
鐮刀龍下目 Therizinosauria
•鑄鐮龍Falcarius
•鐮刀龍超科 Therizinosauroidea
•北票龍Beipiaosaurus
•二連龍Erliansaurus
•死神龍Erlikosaurus
•內蒙古龍Neimongosaurus
•肅州龍Suzhousaurus
•阿拉善龍科Alxasauridae
•鐮刀龍科Therizinosauridae
其他可能是鐮刀龍類的物種包含:鞘虛骨龍、以及早侏羅紀的峨山龍,如果峨山龍屬於鐮刀龍類,那它們將是已知最早的虛骨龍類恐龍。
系統發生學
在1997年,戴爾·羅素(Dale Russell)首次定義鐮刀龍下目演化支為:包含阿拉善龍、死神龍、南雄龍、慢龍、鐮刀龍,以及所有親緣關係接近以上各屬,而離偷蛋龍下目、似鳥龍科、傷齒龍科較遠的所有物種。在2005年,保羅·塞里諾(Paul Sereno)將這定義修正為:包含鐮刀龍,但不包含似鳥龍、偷蛋龍、鳥面龍、暴龍、傷齒龍在內的最大演化支。
在2001年,Zhang等人首次將鐮刀龍超科定義為:獸腳亞目之中,親緣關係接近於鐮刀龍,而離鳥類較遠的所有物種;鐮刀龍超科有效地取代慢龍下目,慢龍下目從未被定義成演化支過。然而,這個定義與較早定義的鐮刀龍下目範圍相同。而Clark在2004年提出一個更為狹隘的定義:鐮刀龍與北票龍的最近共同祖先,以及最近共同祖先的所有後代;這個定義不包含原始的鐮刀龍類,例如鑄鐮龍,但允許鐮刀龍下目繼續包含者鐮刀龍超科。這個定義由Maryanska與巴思缽(2004年)、賽里諾(2005年)以及Zanno(2009年、2010年)所沿用;但其他的研究,例如Senter(2007年)僅使用鐮刀龍超科。
以下分類圖是根據菲力·森特(Phil Senter)在2007年的研究:
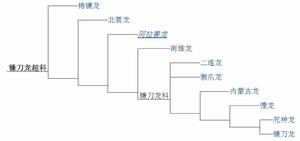 演化樹(2007)
演化樹(2007)以下分類圖來自於Lindsay E. Zanno的2010年鐮刀龍類重新研究:
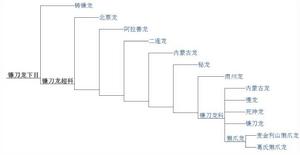 演化樹(2010)
演化樹(2010)以下分類圖來自於Hanyong Pu等人2013年的研究:
古生物學
關於鐮刀龍類的行為模式,過去很少相關研究,但近年發現與研究,開始研究鐮刀龍類的古生物學。現已發現數個蛋巢,內有接近圓形的恐龍蛋。鐮刀龍類的親代可能產下蛋後,用土壤掩埋,然後置之不理。近年研究指出,鐮刀龍類的剛孵化幼年個體,已經生長良好,是早熟性動物,可能不需要太多親代養育。
2012年的一個研究使用電腦斷層掃描,研究死神龍的頭顱骨與顱腔結構,發現它們具有大型前腦,並發現它們腦部的平衡感官、聽覺、嗅覺區域發展良好。研究人員推論,這些特徵有助於鐮刀龍類躲避掠食動物、搜尋食物、或是其他社會行為。
