名稱:氧化磷酸化體
英文:Oxidative phosphorylation body
氧化磷酸化
 氧化磷酸化體
氧化磷酸化體概念:氧化磷酸化(oxidative phosphorylation)是指在生物氧化中伴隨著ATP生成的作用。有代謝物連線的磷酸化和呼吸鏈連線的磷酸化兩種類型。即ATP生成方式有兩種。一種是代謝物脫氫後,分子內部能量重新分布,使無機磷酸酯化先形成一個高能中間代謝物,促使ADP變成ATP。這稱為底物水平磷酸化。
名詞解釋
1. 生物氧化(biological oxidation)
2. 呼吸鏈(respiratory chain)
3. 氧化磷酸化(oxidative phosphorylation)
4. 磷氧比P/O(P/O)
5. 底物水平磷酸化(substrate level phosphorylation)
6. 能荷(energy charge)
氧化磷酸化的概念和偶聯部位
 氧化磷酸化體
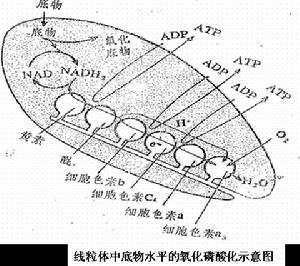
氧化磷酸化體1.概念:氧化磷酸化(oxidative phosphorylation)是指在生物氧化中伴隨著ATP生成的作用。有代謝物連線的磷酸化和呼吸鏈連線的磷酸化兩種類型。即ATP生成方式有兩種。一種是代謝物脫氫後,分子內部能量重新分布,使無機磷酸酯化先形成一個高能中間代謝物,促使ADP變成ATP。這稱為底物水平磷酸化。如3-磷酸甘油醛氧化生成1,3-二磷酸甘油酸,再降解為3-磷酸甘油酸。另一種是在呼吸鏈電子傳遞過程中偶聯ATP的生成。生物體內95%的ATP來自這種方式。
2.偶聯部位:根據實驗測定氧的消耗量與ATP的生成數之間的關係以及計算氧化還原反應中ΔGO'和電極電位差ΔE的關係可以證明。
P/O比值是指代謝物氧化時每消耗1摩爾氧原子所消耗的無機磷原子的摩爾數,即合成ATP的摩爾數。實驗表明, NADH在呼吸鏈被氧化為水時的P/O值約等於3,即生成3分子ATP;FADH2氧化的P/O值約等於2,即生成2分子ATP。
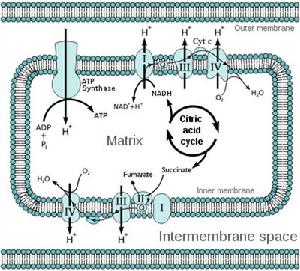
氧-還電勢沿呼吸鏈的變化是每一步自由能變化的量度。根據ΔGO'= - nFΔE O'(n是電子傳遞數,F是法拉第常數),從NADH到Q段電位差約0.36V,從Q到Cytc為0.21V,從aa3到分子氧為0.53V,計算出相應的ΔGO'分別為69.5、40.5、102.3kJ/mol。於是普遍認為下述3個部位就是電子傳遞鏈中產生ATP的部位。
NADH→NADH脫氫酶→‖Q → 細胞色素bc1複合體→‖Cytc →aa3→‖O2
胞液中NADH的氧化
 氧化磷酸化體
氧化磷酸化體糖代謝中的三羧酸循環和脂肪酸β-氧化是線上粒體內生成NADH(還原當量),可立即通過電子傳遞鏈進行氧化磷酸化。在細胞的胞漿中產生的NADH ,如糖酵解生成的NADH則要通過穿梭系統(shuttle system)使NADH的氫進入線粒體內膜氧化。
(一)α-磷酸甘油穿梭作用
這種作用主要存在於腦、骨骼肌中,載體是α-磷酸甘油。
胞液中的NADH在α-磷酸甘油脫氫酶的催化下,使磷酸二羥丙酮還原為α-磷酸甘油,後者通過線粒體內膜,並被內膜上的α-磷酸甘油脫氫酶(以FAD為輔基)催化重新生成磷酸二羥丙酮和FADH2,後者進入琥珀酸氧化呼吸鏈。葡萄糖在這些組織中徹底氧化生成的ATP比其他組織要少,1摩爾G→36摩爾ATP。
(二)蘋果酸-天冬氨酸穿梭作用
主要存在肝和心肌中。1摩爾G→38摩爾ATP
胞液中的NADH在蘋果酸脫氫酶催化下,使草醯乙酸還原成蘋果酸,後者藉助內膜上的α-酮戊二酸載體進入線粒體,又線上粒體內蘋果酸脫氫酶的催化下重新生成草醯乙酸和NADH。NADH進入NADH氧化呼吸鏈,生成3分子ATP。草醯乙酸經穀草轉氨酶催化生成天冬氨酸,後者再經酸性胺基酸載體轉運出線粒體轉變成草醯乙酸。
氧化磷酸化偶在線上制
 氧化磷酸化體
氧化磷酸化體(一)化學滲透假說(chemiosmotic hypothesis)
1961年,英國學者Peter Mitchell提出化學滲透假說(1978年獲諾貝爾化學獎),說明了電子傳遞釋出的能量用於形成一種跨線粒體內膜的質子梯度(H 梯度),這種梯度驅動ATP的合成。這一過程概括如下:
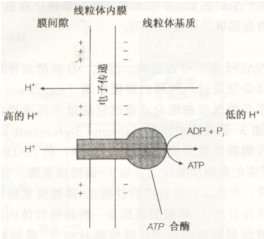
1.NADH的氧化,其電子沿呼吸鏈的傳遞,造成H 被3個H 泵,即NADH脫氫酶、細胞色素bc1複合體和細胞色素氧化酶從線粒體基質跨過內膜泵入膜間隙。
2.H 泵出,在膜間隙產生一高的H 濃度,這不僅使膜外側的pH較內側低(形成pH梯度),而且使原有的外正內負的跨膜電位增高,由此形成的電化學質子梯度成為質子動力,是H 的化學梯度和膜電勢的總和。
3.H 通過ATP合酶流回到線粒體基質,質子動力驅動ATP合酶合成ATP。
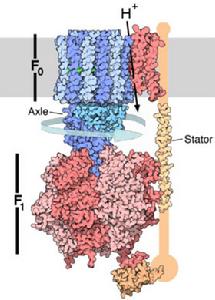
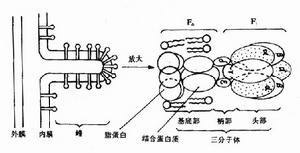
(二)ATP合酶
ATP合酶由兩部分組成(Fo-F1),球狀的頭部F1突向基質液,水溶性。亞單位Fo埋在內膜的底部,是疏水性蛋白,構成H 通道。在生理條件下,H 只能從膜外側流向基質,通道的開關受柄部某種蛋白質的調節。
影響氧化磷酸化的因素
 氧化磷酸化體
氧化磷酸化體(一)抑制劑
能阻斷呼吸鏈某一部位電子傳遞的物質稱為呼吸鏈抑制劑。
魚藤酮、安密妥在NADH脫氫酶處抑制電子傳遞,阻斷NADH的氧化,但FADH2的氧化仍然能進行。
抗黴素A抑制電子在細胞色素bc1複合體處的傳遞。
氰化物、CO、疊氮化物(N3-)抑制細胞色素氧化酶。
對電子傳遞及ADP磷酸化均有抑制作用的物質稱氧化磷酸化抑制劑,如寡黴素。
(二)解偶聯劑
2,4-二硝基苯酚(DNP)和頡氨黴素可解除氧化和磷酸化的偶聯過程,使電子傳遞照常進行而不生成ATP。DNP的作用機制是作為H 的載體將其運回線粒體內部,破壞質子梯度的形成。由電子傳遞產生的能量以熱被釋出。
(三)ADP的調節作用
正常機體氧化磷酸化的速率主要受ADP水平的調節,只有ADP被磷酸化形成ATP,電子才通過呼吸鏈流向氧。如果提供ADP,隨著ADP的濃度下降,電子傳遞進行,ATP在合成,但電子傳遞隨ADP濃度的下降而減緩。此過程稱為呼吸控制,這保證電子流只在需要ATP合成時發生。
氧化磷酸化作用
氧化磷酸化作用是需氧細胞生命活動的基礎,是主要的能量來源。真核細胞是線上粒體內膜上進行。
生物氧化與氧化磷酸化
1.氧化磷酸化作用
高勢能電子從NADH或FADH2沿呼吸鏈傳遞給氧的過程中,所釋放的能量轉移給ADP形成ATP,即ATP的形成與電子傳遞相偶聯,稱為氧化磷酸化作用,其特點是需要氧分子參與。
氧化磷酸化作用與底物水平磷酸化作用是有區別的:底物水平磷酸化作用是指代謝底物由於脫氫或脫水,造成其分子內部能量重新分布,產生的高能鍵所攜帶的能量轉移給ADP生成ATP,即ATP的形成直接與一個代謝中間高能磷酸化合物(如磷酸烯醇式丙酮酸、1,3-二磷酸甘油酸等)上的磷酸基團的轉移相偶聯,其特點是不需要分子氧參加。
2.P/O比和磷酸化部位
磷氧比(P/O)是指一對電子通過呼吸鏈傳遞到氧所產生ATP的分子數。由NADH開始氧化脫氫脫電子,電子經過呼吸鏈傳遞給氧,生成3分子ATP,則P/O比為3。這3分子ATP是在三個部位上生成的,第一個部位是在NADH和COQ之間,第二個部位是在Cytb與Cytc1之間;第三個部位是在Cytaa3和氧之間。如果從FADH2開始氧化脫氫脫電子,電子經過呼吸鏈傳遞給氧,只能生成2分子ATP,其P/O比為2。
(1)氧化磷酸化的解偶聯作用 在完整線粒體內,電子傳遞與磷酸化是緊密偶聯的,當使用某些試劑而導致的電子傳遞與ATP形成這兩個過程分開,只進行電子傳遞而不能形成ATP的作用,稱為解偶聯作用。
(2)氧化磷酸化的解偶聯劑 能引起解偶聯作用的試劑稱為解偶聯劑,解偶聯作用的實質是解偶聯劑消除電子傳遞中所產生的跨膜質子濃度或電位梯度,只有電子傳遞而不產生ATP。
(3)解偶聯劑種類 典型的解偶聯劑是化學物質2,4-二硝基苯酚(DNP),DNP具弱酸性,在不同pH環境可結合H 或釋放H ;並且DNP具脂溶性,能透過磷脂雙分子層,使線粒體內膜外側的H 轉移到內側,從而消除H 梯度。此外,離子載體如由鏈黴素產生的抗菌素——纈氨黴素,具脂溶性,能與K 離子配位結合,使線粒體膜外的K 轉運到膜內而消除跨膜電位梯度。另外還有存在於某些生物細胞線粒體內膜上的天然解偶聯蛋白,該蛋白構成的質子通道可以讓膜外質子經其通道返回膜內而消除跨膜的質子濃度梯度,不能生成ATP而產生熱量使體溫增加。
解偶聯劑與電子傳遞抑制劑是不同的,解偶聯劑只消除內膜兩側質子或電位梯度,不抑制呼吸鏈的電子傳遞,甚至加速電子傳遞,促進呼吸底物和分子氧的消耗,但不形成ATP,只產生熱量。
4.氧化磷酸化的作用機理
與電子傳遞相偶聯的氧化磷酸化作用機理雖研究多年,但仍不清楚。曾有三種假說試圖解釋其機理。這三種假說為:化學偶聯假說、構象偶聯假說、化學滲透假說。
(1)化學偶聯假說 認為電子傳遞中所釋放的自由能以一個高能共價中間物形式暫時存在,隨後裂解將其能量轉給ADP以形成ATP。但不能從呼吸鏈中找到高能中間物的實例。
(2)構象偶聯假說 認為電子沿呼吸鏈傳遞釋放的自由能使線粒體內膜蛋白質發生構象變化而形成一種高能形式暫時存在。這種高能形式將能量轉給F0F1-ATP酶分子使之發生構象變化,F0F1-ATP酶復原時將能量轉給ADP形成ATP。
(3)化學滲透假說 該假說由英國生物化學家Peter Mitchell提出的。他認為電子傳遞的結果將H 從線粒體內膜上的內側“泵”到內膜的外側,於是在內膜內外兩側產生了H 的濃度梯度。即內膜的外側與內膜的內側之間含有一種勢能,該勢能是H 返回內膜內側的一種動力。H 通過F0F1-ATP酶分子上的特殊通道又流回內膜的內側。當H 返回內膜內側時,釋放出自由能的反應和ATP的合成反應相偶聯。該假說目前得到較多人的支持。
實驗證明氧化磷酸化作用的進行需要完全的線粒體內膜存在。當用超音波處理線粒體時,可將線粒體內膜嵴打成片段:有些片段的嵴膜又重新封閉起來形成泡狀體,稱為亞線粒體泡(內膜變為翻轉朝外)。這些亞線粒體泡仍具有進行氧化磷酸化作用的功能。在囊泡的外面可看到F1球狀體。用尿素或胰蛋白酶處理這些囊泡時,內膜上的球體F1脫下,F0留在膜上。這種處理過的囊泡仍具有電子傳遞鏈的功能,但失去合成ATP的功能。當將F1球狀體再加回到只有F0的囊泡時,氧化磷酸化作用又恢復。這一實驗說明線粒體內膜嵴上的酶(F0)起電子傳遞的作用,而其上的F1是形成ATP的重要成分,F0和F1是一種酶的複合體。
5.能荷
細胞中存在三種腺苷酸即AMP、ADP、ATP,稱為腺苷酸庫。在細胞中ATP、ADP和AMP在某一時間的相對數量控制著細胞活動。Atkinson提出了能荷的概念。認為能荷是細胞中高能磷酸狀態的一種數量上的衡量,能荷大小可以說明生物體中ATP-ADP-AMP系統的能量狀態。
能荷=
可看出,能荷的大小決定於ATP和ADP的多少。能荷的從0到1.0,當細胞中都是ATP時,能荷為1.0。此時,可利用的高能磷酸鍵數量最大。都為ADP時,能荷為0.5,系統中有一半的高能磷酸健。都為AMP時,能荷為0,此時無高能磷酸化合物存在。實驗證明能荷高時可抑制ATP的生成,卻促進ATP的利用。也就是說,能荷高可促進合成代謝而抑制分解代謝,相反,能荷低則促進分解代謝而抑制合成代謝。
能荷調節是通過ATP、ADP和AMP分子對某些酶分子進行變構調節來實現的。
5、線粒體的穿梭系統
真核生物在細胞質中進行糖酵解時所生成的NADH是不能直接透過線粒體內膜被氧化的,但是NADH+H 上的質子可以通過一個穿梭的間接途徑而進入電子傳遞鏈。3-磷酸甘油的穿梭過程是最早發現的。其過程是胞質中NADH十H 在3-磷酸甘油脫氫酶作用下與磷酸二羥丙酮反應生成3-磷酸甘油。3-磷酸甘油可進入線粒體,線上粒體內膜上的3-磷酸甘油脫氫酶(輔基為FAD)作用下,生成磷酸二羥丙酮和FADH2。磷酸二羥丙酮透出線粒體,繼續作為氫的受體,FADH2將氫傳遞給CoQ進入呼吸鏈氧化,這樣只能產生2分於ATP。
在動物的肝、腎及心臟的線粒體存在另一種穿梭方式,即草醯乙酸-蘋果酸穿梭。這種方式在胞液及線粒體內的脫氫酶輔酶都是NAD+,所以胞液中的NADH H 到達線粒體內又生成NADH+H 。從能量產生來看,草醯乙酸-蘋果酸穿梭優於α- 磷酸甘油太空梭制;但α-磷酸甘油太空梭制比草醯乙酸-蘋果酸穿梭速度要快很多。
