
分類
原核生物的rRNA分三類:5SrRNA、16SrRNA和23SrRNA。
真核生物的rRNA分四類:5SrRNA、5.8SrRNA、18SrRNA和28SrRNA。
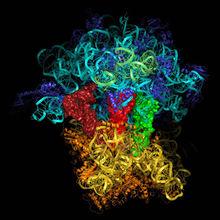 核糖體RNA
核糖體RNAS為大分子物質在超速離心沉降中的一個物理學單位,可間接反映分子量的大小。原核生物和真核生物的核糖體均由大、小兩種亞基組成。
在人基因組的四種rRNA基因中, 18S、5.8S和28S rRNA基因是串聯在一起的,每個基因被間隔區隔開, 5S的rRNA基因則是編碼在另一條染色體上。
特點
核糖體RNA在各種生物中都有其特性,因此可以從不同生物的rRNA的對比中得出關於生物進化歷程的結論。
rRNA為肽醯轉移酶(peptidyl transferase)時,催化使肽鍵形成,不需要額外的能量。
過去認為,大亞基的蛋白質具有酶的活性,促使肽鍵形成,故稱為轉肽酶。20世紀90年代初,H.F.Noller等證明大腸桿菌的23SrRNA能夠催化肽鍵的形成,才證明核糖體是一種核酶,從而根本改變了傳統的觀點。核糖體催化肽鍵合成的是rRNA,蛋白質只是維持rRNA構象,起輔助的作用。
組成
rRNA一般與核糖體蛋白質結合在一起,形成核糖體(ribosome),如果把rRNA從核糖體上除掉,核糖體的結構就會發生塌陷。原核生物的核糖體所含的rRNA有5S、16S及23S三種。S為沉降係數(sedimentation coefficient),當用超速離心測定一個粒子的沉澱速度時,此速度與粒子的大小直徑成比例。5S含有120個核苷酸,16S含有1540個核苷酸,而23S含有2900個核苷酸。而真核生物有4種rRNA,它們分子大小分別是5S、5.8S、18S和28S,分別具有大約120、160、1900和4700個核苷酸。
rRNA是單鏈,它包含不等量的A與U、G與C,但是有廣泛的雙鏈區域。在雙鏈區,鹼基因氫鍵相連,表現為髮夾式螺旋。
rRNA在蛋白質合成中的功能尚未完全明了。但16 S的rRNA3’端有一段核苷酸序列與mRNA的前導序列是互補的,這可能有助於mRNA與核糖體的結合。
結構
 核糖體RNA
核糖體RNA測定rRNA的空間排列方式的方法主要有電鏡法和交聯法。其功能部位通過幾種方法確定在70S核糖體圖中顯示了rRNA分子的結合部位和方向。在電鏡下,16SrRNA的排列呈V型,一個臂比一個臂稍厚和長。23S的大小和形狀可與50S"皇冠"式樣很好匹配。有結論認為,rRNA形成了核糖體亞基的骨架,蛋白質與其結合。一般來說,rRNA骨架不發生大的構象改變。用免疫電鏡法已確定在亞基內rRNA的某些特徵。使用抗N6,6-二甲基腺苷(位於16SrRNA3'末端24和25位)抗體,確定了修飾鹼基區段(指16SrRNA3'端約25個鹼基)位於30S亞基頭和體之間。16SrRNA的第526位的m7G處於30S上1/3和下2/3交界處。16S、5S和23SrRNA的內部交聯已被研究。證明在5SrRNA內G41和G72交聯,這種交聯屬三級結構反應,利用此反應已經構建了一處改進的5SrRNA分子三維模型。此外,RNA-蛋白質交聯研究也是測定亞基內rRNA分子空間排列的非常有用的方法。
一般認為,核糖體的基本功能依賴於其中的rRNA,核糖體蛋白質起著加強rRNA功能的作用。核糖體最初由rRNA構建,在進化過程中一些蛋白質加在其上。在體內外的實驗均證明了缺乏某些蛋白質的核糖仍有生物活性;此外,rRNA基因(rDNA)突變及甲基化等均可引起對抗菌素(如紅黴素、氯黴素)的抵抗。
功能
在核糖體中,rRNA是起主要作用的結構成分,是結構和功能核心,主要功能是:
(1)具有肽醯轉移酶的活性。
(2)為tRNA提供結合位點。
(3)為多種蛋白質合成因子提供結合位點。
(4)在蛋白質合成起始時,參與同mRNA選擇性的結合以及在肽鏈的延伸中與mRNA結合。
(5)此外,核糖體大小亞單位的結合、校正閱讀、無意義鏈或框架漂移的校正以及抗生素的作用等都與rRNA有關。
