 花粉管
花粉管花粉管,是萌發的花粉粒內壁突出,花粉粒的內壁通過花粉外壁上的萌發孔(或溝)向外伸出的細管,從萌發孔伸出而形成的管狀結構。主要作用是將其攜帶的精子和其他內容物運至卵器或卵細胞內,以利於受精作用。花粉管是雄配子體的一部分。在被子植物中,雌配子體(胚囊)深藏於雌蕊子房的胚珠內,含有精細胞的花粉粒(雄配子體)必須經過萌發,形成花粉管,並通過花粉管將精細胞送入胚囊,才能使兩性細胞相遇而結合,完成受精過程。 花粉管是研究細胞極性生長的好材料之一,也是研究細胞骨架和囊泡運輸的理想體系。利用植物在開花、受精過程中形成的花粉管通道,將外源DNA導入花粉管受精卵細胞,並進一步地被整合到受體細胞的基因組中,隨著受精卵的發育而成為帶轉基因的新個體。
花粉管的形成
被子植物的花粉粒萌發時,營養細胞內壓增加而迫使花粉粒內壁向著萌發孔突出,形成一個(或幾個)花粉管。
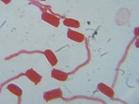 花粉管
花粉管花粉管的生長
花粉管生長的細胞生化基礎
 花粉管
花粉管花粉管是研究細胞極性生長的好材料之一,也是研究細胞骨架和囊泡運輸的理想體系。清華大學李一勤研究組通過 DNA 微陣列法從百合 (Lilium longiflorum) 中分離到一個在花粉管高度表達的基因 LIANK,LIANK(編碼一個含有5個 ANKYRIN 重複結構域的 RING 鋅指蛋白。LIANK 蛋白定位在細胞膜泡上,在體外具有泛素連線酶活性。LIANK 瞬時過表達和 RNAi 都影響百合花粉管的正常生長,說明 LIANK 參與了花粉管極性生長,但具體機理還有待進一步研究(Huang et al.,2006)。中國科學院植物研究所林金星研究組發現蛋白酶體抑制劑 MG132 和 Epoxomicin 抑制花粉管生長和改變花粉管形態。進一步分析發現抑制劑處理引起內質網液泡化並積累泛素化的蛋白,同時細胞骨架系統也受到破壞,但對鈣離子梯度影響不大,其結果是果膠和纖維素等細胞壁物質不能運輸到快速生長的花粉管頂部,導致花粉管生長受阻(Sheng et al.,2006)。以上研究結果為了解泛素/蛋白酶體途徑在花粉管中的作用機制提供了細胞學證據。
新技術手段對植物科學研究起了極大的促進作用。近來,全內反射螢光顯微技術的興起,實現了對單個螢光分子的直接探測,用全內反射產生的隱失波可使照明區域限定在樣品表面的一薄層範圍內,對於觀察膜泡類細胞器尤為有效。林金星研究組探索了用消散波顯微技術研究花粉管極性生長過程中膜泡的動態變化 (Wang et al.,2006d),發現膜泡呈現複雜的震盪現象,而不是以前認為的簡單布朗運動,肌動蛋白細胞骨架對膜泡動態影響比微管骨架大。同樣,利用蛋白組學手段,林金星研究組發現簡單的肌動蛋白聚合抑制劑 Latrunculin B 處理,會引起細胞器和蛋白組學水平的巨大變化 (Chen et al.,2006f)。這些工作加深了我們對花粉管極性生長的認識。
採用花粉液體培養法研究了植物生長調節物質對梨花粉萌發和花粉管生長的影響,結果表明:較低濃度的赤黴素、三十烷醇、吲哚乙酸及2,4-D均能促進花粉萌發和花粉管生長,而超過一定濃度時卻起抑制作用,最適宜於花粉萌發和花粉管生長的赤黴素濃度為50~300mg/L,三十烷醇為3~100mg/L,吲哚乙酸為5~25mg/L,2,4-D為5~10mg/L。萘乙酸對花粉萌發和花粉管生長有抑制作用,抑制程度隨培養基內其濃度的增大而加強。多效唑和脫落酸對花粉萌發有抑制作用,其抑制程度隨濃度的上升而增強,但他們對花粉管生長卻有促進怍用,其最適宜於花粉管生長的濃度分別為400mg/L和60mg/L,超過此濃度後,促進作用又有所下降,甚至出現抑制作用,如多效唑濃度達到1000mg/L時,能強烈地抑制花粉管生長。
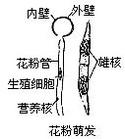 花粉管
花粉管鶴頂蘭花粉管在子房中的生長途徑
運用掃描電鏡對鶴頂蘭(Phaius tankervilliae(Aiton)B1)花粉管在子房內的生長途徑進行了觀察.結果表明:花粉管在子房中的生長途徑可以分為3個階段:(1)沿子房壁軸向生長階段,從授粉開始至大孢子母細胞四分體時期,花粉管經過合蕊柱到達子房,經由胎座基部沿子房壁軸向生長;(2)沿子房徑向生長階段,二核胚囊之後,花粉管在胚珠之間穿梭,以徑向生長為主;(3)朝珠孔定向生長階段,胚囊成熟時,花粉管朝珠孔定向生長進入胚囊.實驗結果說明花粉管的定向生長受胚珠的分子信號調控。
花粉管作用
花粉粒在雌蕊柱頭上萌發時,其內壁經外壁上的萌發孔向外突出所形成的管狀物,即是花粉管,它具有頂端生長的特性。在其生長過程中,花粉粒內的營養細胞和生殖細胞(二細胞花粉)或營養細胞與生殖細胞分裂形成的兩個精子(三細胞花粉),相繼進入花粉管。在花粉管中,二細胞花粉的生殖細胞核完成分裂,形成2個精子。花粉管穿過柱頭,經花柱進入子房,最後準確地伸向胚珠和胚囊,將花粉管中的兩個精子和全部內含物釋放到胚囊中,使受精過程得以完成。花粉管在花柱中生長,除利用花粉粒本身貯存的物質外,也從花柱組織中吸收營養,供生長和建造管壁之用。柱頭、花柱、子房內壁、胚珠、胎座等都有引導花粉管朝向胚珠定向生長的作用。一般一個花粉粒只長出一個花粉管。具多萌發孔的花粉粒,可以同時長出幾個花粉管(如錦葵科、葫蘆科),但最終只有一個能繼續生長。在柱頭上萌發的花粉粒往往不止一個,故在花柱中生長的花粉管常為多個,但最終能進到胚囊的通常只有一個。花粉管的生長速度差異很大。例如,秋水仙的花粉管須經6個月的時間,才能貫穿花柱;番紅花屬植物的花粉管,在2~3日內,即可穿過10厘米長的花柱到達子房;而橡膠草只需半小時左右的時間,即可完成傳粉到受精的整個過程。花粉管的長短因植物花柱長短而異,通常花柱長的,花粉管也長,花柱短的,花粉管也短。
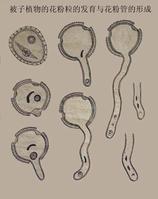 花粉管
花粉管種子植物生殖器官的結構相當複雜,在有性生殖過程中產生了花粉管,藉助花粉管,受精不再依賴水的條件,這是種子植物適應陸地生活的一個重要因素。產生雌配子的雌配子體在大孢子囊即胚珠的珠心處發育。裸子植物中的大多數仍保留頸卵器的結構,卵在頸卵器內形成。在小孢子囊中發育的花粉粒,即前期的雄配子體。在雄配子體中形成兩個精子。精子多不具鞭毛,但在蘇鐵目和銀杏目仍具遊動的精子。被子植物的雌配子體稱為胚囊,卵直接在其中產生;精子完全失去鞭毛。這些特點決定了種子植物的受精必先經過傳粉。裸子植物的胚珠是裸露的,經過傳粉,花粉被送到胚珠的珠孔處,而在被子植物中,胚珠著生在雌蕊的子房內,花粉粒不能直達胚珠而是落在柱頭上,再由萌發生長的花粉管將精子輸送到雌配子附近。在裸子植物中,進入頸卵器的兩個精子只有一個與卵融合。但在被子植物中進入胚囊的兩個精子則分別與卵和極核融合。這就是被子植物特有的。裸子植物的雄配子即花粉粒,通常由風力傳播,經珠孔直接進到胚珠,在珠心上萌發,形成花粉管,進入胚囊,將由生殖細胞所產生的2個精子直接送到頸卵器內,其中1個具有功能的精子和卵細胞結合,完成受精作用,因此受精不在受水的限制。雄配子體是有小孢子發育成的花粉粒,在多數種類僅有4細胞組成:2個退化的原葉細胞,1個生殖細胞和1個管細胞。雌配子體由大孢子發育而來,在近珠孔端產生2至多個頸卵器,但結構簡短,埋藏於胚囊中,僅有2-4個頸細胞露在外面,頸卵器內有1個卵細胞和1個腹溝細胞,無頸溝細胞,比蕨類植物頸卵器更加退化。雌、雄配子均無獨立生活能力,完全寄生在孢子體上。
受精
受精作用
雌雄配子即卵子和精子的融合過程稱為受精作用(fertilization)。在被子植物中,雌配子體(胚囊)深藏於雌蕊子房的胚珠內,含有精細胞的花粉粒(雄配子體)必須經過萌發,形成花粉管,並通過花粉管將精細胞送入胚囊,才能使兩性細胞相遇而結合,完成受精過程。
傳粉完成後,落在柱頭上的花粉粒被柱頭分泌的粘液所粘住,以後花粉內壁在萌發孔處向外突出,並繼續伸長,形成花粉管,這一過程即為花粉粒的萌發。花粉管形成後能繼續向下引伸,先穿過柱頭,然後經花柱而達子房,同時,花粉粒細胞的內含物全部注入花粉管中,並向花粉管頂端集中,此前,花粉粒的發育由單核細胞分裂形成2個細胞,一個是營養細胞,另一個是生殖細胞,生殖細胞在移入花粉管後再分裂形成2個精子,也有少數植物的生殖細胞在進入花粉管前就已分裂成2個精子。花粉管進入子房後,一般通過珠孔進入胚囊(也有經過合點進入胚囊的),此時花粉管先端破裂,兩個精子進入胚囊(這時營養細胞大多已解體消失),其中一個精子與卵結合成合子,將來發育成種子的胚,另一個精子與極核結合而發育成種子的胚乳。
 花粉管
花粉管卵細胞和極核同時和兩個精子分別完成融合的過程,是被子植物有性生殖的特有現象,稱為雙受精(double fertilization),它在融合雙親遺傳特性,加強後代個體的生活力和適應性方面具有重要意義,是植物界有性生殖過程中最進化、最高級的形式。
雙受精過程
被子植物的雄配子體形成的兩個精子,一個與卵融合形成二倍體的合子,另一個與中央細胞的極核(通常兩個)融合形成初生胚乳核的現象。雙受精後由合子發育成胚,初生胚乳核發育成胚乳。
被子植物胚囊中極核同卵一樣受精產生了具有父本和母本遺傳性的通常是三倍體的胚乳,由這種胚乳“哺育”胚可能使後代更加鞏固它雙親的特性並更富有生命力,因此有人認為雙受精是被子植物繁盛的一個重要原因。
配子配合前的階段可見:①花粉萌發和花粉管在花柱中生長。花粉由各種媒介傳到雌蕊的柱頭上後,立即與雌蕊發生相互作用。在親和的情況下,花粉萌髮長出花粉管並鑽入柱頭。花粉管進入柱頭後,繼續在花柱中生長。花粉管依賴其末端生長,當生長到一定長度後,原來在花粉中含有的物質全部集中到花粉管的前端。在生長過程中花粉管除利用自身的貯藏物質外,同時也可能從花柱中吸收營養。②花粉管進入胚囊和釋放內容物。花粉管伸入子房後,沿子房內壁或胎座繼續生長,直達胚珠,經珠孔進入胚珠,最後到達胚囊。花粉管沿一定道路的生長,一般認為是受助細胞分泌的化學物質的吸引。
通常把精子與卵的融合稱為配子配合,而精子與極核的融合稱為三核併合。這兩種融合是差不多同時發生的。1973年,美國植物胚胎學家W.A.詹森提出了被子植物雙受精作用中精子轉移至卵細胞和中央細胞的模式圖。精子先釋放至退化的助細胞(質膜已經消失),由於花粉管內容物釋放時的力量,兩個精子被分別轉移至卵細胞的質膜和中央細胞的質膜處並與之接觸,接觸處的質膜溶解,兩個精核分別轉移至卵和中央細胞的細胞質中。按詹森的精子入卵模式,只是精子核轉移至雌性細胞中。實際上在被子植物的受精作用中,精子的細胞質是否參加迄今尚無定論。被子植物雄核與雌核融合的基本程式是:①雄性核與雌性核接觸。②雄性核和雌性核之間的核膜融合。③精子核的染色質在卵核內分散和出現雄性核仁。④雄性核的染色質與雌性核的染色質混合。⑤雄性和雌性的核仁融合為一個大的核仁。精核同極核(或次生核)的融合過程與精核同卵核的融合程式基本相同。
 花粉管
花粉管從花粉落到柱頭上至雄性和雌性核完全融合所經歷的時間,因植物不同而異。間隔期短的只有十幾分鐘,長的可達一年。大多數植物為幾小時至48小時之間。精卵融合與精子同極核融合通常是同時發生的,也可能精子同極核的融合更早發生,而且一般更快完成融合和進入分裂。這樣胚乳的發育早於胚,可以為胚的發育提供必需的營養物質。傳粉至受精間隔期的長短還受環境因素,特別是溫度的影響。溫度對花粉萌發和花粉管的生長速度有直接的影響。
在正常的情況下,一個胚囊只接受兩個精子,但是有時由於不止一個花粉管進入或是由於一個花粉管中形成不止一對精子,於是胚囊中出現有多於一對精子的現象。胚囊中額外精子的存在將有可能引起下面兩種異常受精作用:①多精入卵。有兩種不同的情況:一種是進入卵中多餘的精子在卵中退化,另一種是附加的精子也與卵核融合,這種異常的受精形式只在極少數植物中有細胞學的證據。②額外的精子與胚囊中的其他細胞受精。這種現象曾在一種慈菇屬植物報導過。這種植物的助細胞形態上與卵細胞相似,有兩個花粉管進入胚囊並放出額外的兩個精子,在受精後的胚囊中有時看到3個原胚存在,並有兩個花粉管在胚囊的上部。因此推測助細胞與進入的額外精子受精而產生了多胚。
此外,在受精過程中,助細胞和反足細胞均破壞消失。花經過受精後,胚珠發育成種子,子房發育成果實。
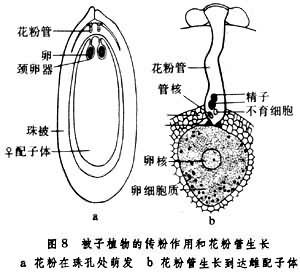 花粉管
花粉管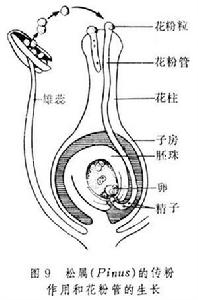 花粉管
花粉管花粉管的技術運用
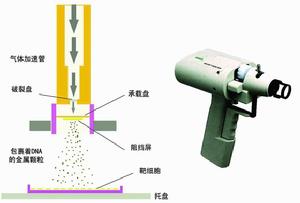
花粉管通道法
在授粉後向子房注射含目的基因的DNA溶液,利用植物在開花、受精過程中形成的花粉管通道,將外源DNA導入
 花粉管
花粉管利用適用於禾穀類作物的表達載體pGU4ABBar ,採用花粉管通道法 ,將人工合成的蘇雲金芽孢桿菌殺蟲蛋白基因cryIa導入了優良冬小麥品系西農 2 2 0 8和西農 132 ,經PCR和Southernblot鑑定 ,證明獲得了 2 7株導入了cryIa基因的轉基因植株 ,轉化率約為 1.13%~ 1.2 1% ,並通過Westernblot鑑定檢測到了目的蛋白的表達。
利用花粉通道法導入HMW-GSlBx14基因的技術體系已初步建立,轉化受體授粉時間在30一60min之間轉化效果最佳,質粒濃度以300ng/μl為宜;轉化率高達12﹪。
花粉管染色
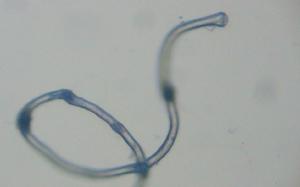 花粉管
花粉管在金魚草和(或)菸草上試驗了三種螢光染料對花粉管進行螢光活體染色與標記的效果。花粉管在異硫氰酸螢光素(FITC,12.5μg/ml)中染色5—6小時,原生質呈綠黃色螢光;換入無染料的培養基後可繼續生長並保持螢光標記,但後期生長受抑。(RB,10μg/nl)染色的效果與上相近,花粉管呈紅色螢光;換入無染料培養基後生長正常,唯後期螢光減弱。螢光素二醋酸酯(FDA,10μg/ml)染花粉粒40分鐘,換入無染料培養基後正常萌發與生長,花粉管原生質呈明亮的綠色螢光。FDA方法具有染色時間短,螢光明亮,兼有活染與生活力鑑定雙重功效等優點。
相關連線
花粉 pollen
 花粉管
花粉管種子植物特有的結構,相當於一個小孢子和由它發育的前期雄配子體。在被子植物成熟花粉粒中包含2個或3個細胞,即一個營養細胞和一個生殖細胞或由其分裂產生的兩個精子。在兩個細胞的花粉粒中,兩個精子是在傳粉後在花粉管中由生殖細胞分裂形成的。在裸子植物的成熟花粉粒中包含的細胞數目變化較大,從1~5個或更多個細胞,其中有1~2個原葉細胞,是雄配子體中殘留的幾個營養細胞,形成後往往隨即退化,在被子植物的雄配子體中已完全消失。
形態 各類植物的花粉各不相同。根據花粉形狀大小,對稱性和極性,萌發孔的數目、結構和位置,壁的結構以及表面雕紋等,往往可以鑑定到科和屬,甚至可以鑑定到植物的種。花粉形態的研究可為分類鑑定和花粉分析中鑑定化石花粉提供依據,同時也為植物系統發育的研究提供有價值的資料。
大多數花粉成熟時分散,成為單粒花粉。但也有兩粒以上花粉粘合在一起的,稱為複合花粉粒。許多花粉結合在一起,在一個藥室中至少有兩塊以上的,稱為花粉小塊。在一個或幾個藥室中全部花粉粒粘合在一起的,稱為花粉塊。花粉小塊和花粉塊主要見於蘭科和蘿藦科植物。
花粉粒在四分體中朝內的部分,稱為近極面。朝外的部分稱為遠極面。連線花粉近極面中心點與遠極面中心的假想中的一條線,稱為極軸,與極軸成直角相交的一條線稱為赤道軸,沿花粉兩極之間表面的中線為赤道。在有極性的花粉中,可以分為等極的,亞等極的和異極的3個類型。花粉通常是對稱的,有兩種不同的對稱性:輻射對稱和左右對稱。
花粉多為球形,赤道軸長於極軸的稱為扁球形;特別扁的稱為超扁球形;相反地,極軸長於赤道軸的稱為長球形,特別長的稱為超長球形。花粉在極面觀所見赤道輪廓,可呈圓形,具角狀,具裂片狀等等。在赤道面觀,花粉輪廓可呈圓形、橢圓形、菱形、方形等等。
花粉大小因種而不同,變化很大。最小的花粉見於紫草科的勿忘草,約(4~8)微米×(2~4)微米。大型花粉直徑為100~200微米〔姜屬〕,120~150微米〔錦葵科的許多屬種,以及牽牛,芭蕉屬等〕。大多數花粉最大直徑約為20~50微米。水生植物大葉藻花粉細長,約為(1200~2900)微米×(3.5~9.5)微米。
萌發孔為花粉壁上變薄的區域,花粉萌發時花粉管往往由萌發孔伸出。萌發孔按其長和寬的比例,通常分為溝、孔兩類。凡長與寬之比大於2的為溝,不到2的為孔。有時短溝和長孔之間不易區分。只具溝或孔的為簡單萌發孔,溝和孔共同組成的為複合萌發孔。萌發孔分布在極面,赤道面或散布於球面。分布於遠極面上的單溝,又稱為槽。萌發孔有許多變異,也有沒有萌發孔的花粉。
花粉壁通常分為兩層,即外壁和內壁。內壁的成分主要是果膠纖維素,抗性較差、在地表容易腐爛,經酸鹼處理則分解;而外壁主要成分是孢粉素,抗腐蝕及抗酸鹼性能強,在地層中經千百萬年仍保持完好,所以研究花粉形態,主要依據外壁的結構。外壁又可分兩層,即外層和內層。外層一般由3層組成,最外層為覆蓋層,其上或具穿孔,發育不完全時,為具半覆蓋層的或無覆蓋層的花粉。下面一層為柱狀層,具有柱狀(或棒狀)結構。再下面一層為基層。
花粉表面光滑或具各種各樣的紋飾(雕紋)。紋飾的類型因種屬而不同。主要的雕紋有顆粒狀,瘤狀,棍棒狀,刺狀,條紋狀,皺波狀,網狀,腦皺狀等等。
生活力和儲藏 成熟花粉的生活力因不同種類而變化很大。許多果樹花粉的生活力,在實驗室可以保持幾個月。禾本科和菊科花粉,往往幾分鐘到幾十分鐘就失去生活力,其他大多數植物花粉的生活力介乎這兩個極端之間。環境因子,特別是溫度與濕度,對花粉的生活力有很大影響。
花粉可以方便地儲藏於膠囊中或小玻璃管內,容器一般置於乾燥器中,以控制濕度(氯化鈣,碳酸鉀或濃硫酸)。最適溫度和濕度因植物種類而不同。禾本科的花粉壽命很短,儲藏比較困難。困難之一是由於它們在水合條件下散粉,而乾燥對它們有害。也不能在冰點下的溫度保存,因為冰凍對花粉生存有害。採集花粉時的溫度和濕度條件,對花粉的生活力也很有關係。
 花粉管
花粉管萌發 在自然界柱頭為花粉萌發提供一個合適的場所。花粉粒落在柱頭上後,即發生吸漲水合,大量吸水,並由於營養核中mRNA的大量形成而產生專一性蛋白質,使花粉萌發,長出花粉管。這些專一性蛋白質與柱頭表膜的專一性蛋白質起識別作用,雙方是否親和,決定於花粉外壁和柱頭表膜上的酶和抗體的特性。如果是親和的,花粉可以萌發,並長進花柱組織和胚囊。許多植物的花粉可以在培養基上萌發,長出花粉管。
花粉萌發一般要求有一種碳水化合物,最常用的是蔗糖。糖在培養基中有兩個作用:一是保持滲透壓,二是作為花粉代謝的底物。硼對花粉管生長有促進作用,鈣對花粉管生長也有明顯作用。
花粉萌發和花粉管生長一般分為四個階段:吸漲、停滯、花粉管發端及花粉管迅速伸長階段。各個階段所需時間因種而不同,並決定於花粉本身所儲存食物及外界因子。花粉管的生長區只限於末端幾個微米。區內富含RNA,蛋白質以及PAS-正反應物質。生長區有許多小泡,可能從高爾基體的嵴膜末端形成。生長區後面的細胞質含細胞器和澱粉體。萌發的特徵是靜止的高爾基器轉變為一個活動的器官,產生小泡,在細胞質中形成液泡。
花粉管核 pollen-tube nucleus
指花粉管內的營養核。幼齡花粉為單核,但由減數分裂所形成的花粉四分體的核在達到成熟花粉為止的過程中進行有絲分裂,母校分化為生殖細胞和營養核。其後因種類的不同,也有營養核退化的情況,但大多數轉向為花粉管核。營養核即花粉管核,大而無定形,生殖細胞稍小,為紡錘形。過去認為花粉管核位於花粉管的頂端,並與花粉管的營養有關,現在已被否定。榆屬(Ulmus)、千里光屬(Senecio)、黃鶴菜屬(Youngia)是在核轉移到花粉管中之前退化。花粉管核變得異常細長的例子也不少,在香堇(Viola odorata),寬度和長度的比為 1∶1,苦 草( Vallisneria asiatica)為1∶27。紫蘇屬(Perilla)、菸草屬(Nicotiana)則有的核成為線狀。歸根到底,花粉管核為痕跡性構造,對花粉管成長並無重大作用。
授粉
授粉是被子植物結成果實必經的過程。花朵中通常都有一些黃色的粉,這叫做花粉。這些花粉需要被傳給同類植物某些花朵。花粉的傳遞過程叫做授粉。
一、根據植物的授粉對象不同,可分為自花授粉(self-pollination)和異花授粉(cross-pollination)兩類。
1、自花傳粉:植物成熟的花粉粒傳到同一朵花的柱頭上,並能正常地受精結實的過程稱自花傳粉 。生產上常把同株異花間和同品種異株間的傳粉也認為是自花傳粉。
能進行自花傳粉的植物稱自花傳粉植物,如水稻、小麥、棉花和桃等,豌豆和花生在花尚未開放,花蕾中的成熟花粉粒就直接在花粉囊中萌發形成花粉管,把精子送入胚囊中受精,這種傳粉方式是典型的自花傳粉,稱閉花受精。
2、異花傳粉:一般情況下既使是兩性花,同一朵花的雌雄蕊也不會一起成熟,因而,一般花的雌蕊接受的花粉是另一朵花的花粉,這就是異花傳粉。當然,雌雄異株植物,雌雄同株中開單性花的,就只有進行異花傳粉了。
油菜、向日葵、蘋果等是異花傳粉的植物。
 花粉管
花粉管二、根據植物的授粉方式不同,可分為自然授粉和人工輔助授粉兩類
1、自然授粉
又分為風媒、蟲媒、水媒、鳥媒等。
風媒: 靠風力傳送花粉的傳粉方式稱風媒(anemophily),藉助這類方式傳粉的花,稱風媒花(anemophilous flower)。大部分禾本科植物和木本植物中的櫟、楊、樺木等都是風媒植物。
蟲媒: 靠昆蟲為媒介進行傳粉方式的稱蟲媒(entomophily),藉助這類方式傳粉的花,稱蟲媒花(entomophilous flower)。多數有花植物是依靠昆蟲傳粉的,常見的傳粉昆蟲有蜂類、蝶類、蛾類、蠅類等。蟲媒花多具一下特點:
(1)多具特殊氣味以吸引昆蟲;
(2)多半能產蜜汁;
(3)花大而顯著,並有各種鮮艷顏色;
(4)結構上常和傳粉的昆蟲形成互為適應的關係。
如馬兜鈴和鼠尾草。
水媒:水生被子植物中的金魚藻、黑藻、水鱉等都是借水力來傳粉的,這類傳粉方式稱水媒(hydrophily)。
鳥媒:其他如借鳥類傳粉的稱鳥媒(ornithophily),傳粉的是一些小形的蜂鳥(Heliothrix au-rita),頭部有長喙,在攝取花蜜時把花粉傳開。蝸牛、蝙蝠等小動物也能傳粉,但不常見。
2、人工輔助授粉
 | |
| 人工輔助受粉 | |
也簡稱人式授粉。農業生產上常採用人工輔助授粉的方法,以克服因條件不足而使傳粉得不到保證的缺陷,以達到預期的產量。在品種復壯的工作中,也需要採取人工輔助授粉,以達到預期的目的。人工輔助授粉可以大量增加柱頭上的花粉粒,使花粉粒所含的激素相對總量有所增加,酶的反應也相應有了加強,起到促進花粉萌發和花粉管生長的作用,受精率可以得到很大提高。
授粉後,花粉粒在柱頭上萌發。隨著花粉管的伸長,營養核與精核進入胚囊內。隨後1個精核與卵細胞受精結合成合子,將來發育為胚(2n)。另1個精核與2個極核受精結合為胚乳核(3n),將來發育成胚乳(3n),故這一過程被稱為雙受精(double fertilization)。通過隨後雙受精最後發育成種子。
英文解釋:
Pollination:
Before seeds can develop inside a flower, pollen grains from the anthers must reach the stigma of the pistil. This process is called pollination.
There are two ways of pollination. in selfpollination, both male and female gametes come from the same plant. In most angiosperms, however, crosspollination brings together gametes from two different parent plants. This means that the pollen from one flower is transferred to a flower on a different plant.
註:圖為給萬壽菊進行人工輔助授粉。
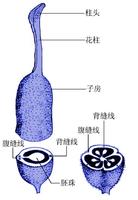
花柱(style)
是柱頭和子房間的連線部分,也是花粉管進入子房的通道。花柱多為細長的結構,如玉米的花柱細長如絲;也有極短不明顯的,如蓮、罌粟。
 花粉管
花粉管有些植物的花柱生於縱向分裂的子房基部,稱花柱基生(gynobasic),如黃芩、丹參等唇形科植物和新疆紫草、紫草等紫草科植物。在蘭科與蘿藦科植物中,雄蕊與花柱合生形成特殊的合蕊柱(gynostemium)。
雌蕊中連線柱頭與子房的部分。一般為細長管狀,也有極短的,如水稻、小麥。花柱是花粉進入子房的通道,花柱的內部結構有兩種類型:開放型(中空的)和閉合型(實心的)。開放型花柱中有寬或窄的花柱道,如百合。其結構是在花柱道周圍常有一層特殊的內表皮細胞,稱為通道細胞,是分泌細胞,能向花柱道分泌物質;閉合型花柱中沒有花柱道,由引導組織充滿。引導組織細胞長形,壁薄,細胞內含豐富的細胞器,細胞間逐漸形成大的胞間隙,並在胞間隙中充滿基質(碳水化物性物質)。不論哪種花柱,都能為花粉管的生長提供營養和某些化學物質,引導花粉管向下生長。花柱內有維管束連線子房、花托和柱頭。
銅肥
開放分類: 金屬、農業、無機化學、植物學、肥料
copper fertilizer
具有銅標明量以提供植物養分為主要功效的物料。
五水硫酸銅是最主要的銅肥。一水硫酸銅、鹼式碳酸銅、氯化銅、氧化銅、氧化亞銅、矽酸銨銅、硫化銅、銅燒結體、銅礦渣、螯合銅等均可作為銅肥施用。
銅是植物必需營養元素之一。銅以陽離子(Cu2+)的形態被植物吸收。
銅的作用:(1)植物體內多種氧化酶的成分,與植物體內的氧化還原反應和呼吸作用有關;(2)對蛋白質代謝及葉綠素的形成有重大影響;(3)能增強光合作用和促進花粉萌發和花粉管伸長,提高結實率。植物正常的含銅量為5~20mg/kg。中國南方長期淹水的水稻土、有機質含量較高的泥炭土、沼澤土以及丘陵坡地的砂質土壤易缺銅。在缺銅土壤上施銅,水稻增產10%~20%。水溶性銅肥如硫酸銅、氯化銅可用作基肥、拌種、浸種,其他銅肥只適於作基肥,銅肥用量過多時,易毒害作物,需慎用。推薦用量:每畝(1畝=667m2)0.5~1kg硫酸銅摻細乾土作基肥。葉面噴施濃度在0.02%以下。
