 耳蝸微音器效應
耳蝸微音器效應耳蝸微音器效應又稱為微音器效應、維弗·布雷(Wever-Bray)效應等,當聲音刺激時,從內耳耳蝸引導出的,與刺激聲振動頻率和波形甚為一致的電位變化,亦即耳蝸有與微音器相似的作用,1930年由維弗(E.G.Wever)和布雷(C.W.Bray)在貓身上發現。
概述
 耳蝸微音器效應
耳蝸微音器效應耳蝸微音器效應最初雖曾與聽神經動作電位相混淆,但以後艾德里安(E.D.Adrian,1931)曾指出兩者是不同的,可明確地加以分離(H.Davis等,1934),即耳蝸微音器電位對阻斷血流、疲勞、寒冷、麻醉劑等比聽神經動作電位有更強的抵抗力,無不應期,即使對高音也能很好地同步,潛伏期短,有的短到0.1毫秒(神經為0.7毫秒)。當把電極置於圓窗處時反應較易證明,其大小達1毫伏。如果把微電極插入耳蝸螺鏇中,
對導出的反應聲閾值進行測定,發現越向頂部對低音越敏感。越近底部對高音越敏感,與赫爾姆霍茨以來的部位說的假設十分一致。對耳蝸微音器效應發生機制的解釋是:伴隨著聲音刺激基底膜發生振動,由於這種振動,基底膜上排列著的毛細胞與被蓋膜接觸,此時因細胞的形變而產生電現象。這一作用可用來確定其他動物的可聽振動頻率。從魚類的球囊和側線器已證明有與耳蝸微音器效應相當的電位變化(Y.Zotterman,1943)。對毛細胞的電現象除在耳蝸外也已在水生動物體表的側線器(一般認為與耳蝸屬相同類型)上進行了研究。毛細胞的毛中有一根運動毛和數十根不動毛,運動毛的運動方向與膜電位之間有一定的關係,向一方向顯示去極化,向另一方向顯示超極化。由於這種膜電位的變化,在毛細胞底部的突觸區遞質向神經末梢釋放,在神經末梢處產生突觸後電位之後開始放電。若從外部觀察膜電位的這一變化,與微音器效應相似,因此這一電位變化稱為耳蝸的微音器效應。耳蝸內淋巴與外淋巴相比,K+的量比Na+要多得多,而接近於K+等滲液;同時在內、外淋巴間可測得+100毫伏的電位差。這一電位差的源基於耳蝸管側壁血管區K+的主動運轉。一般認為,這樣高濃度的K+和內淋巴的正電位對毛細胞的活動和微音器電位的發生起著重要的作用。
作用
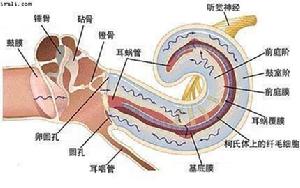
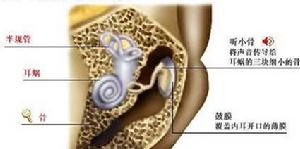
 耳蝸效應
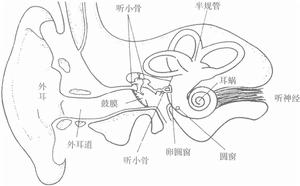
耳蝸效應耳蝸是位聽器官內耳骨迷路的一個組成部分,是傳導並感受聲波的結構。耳蝸是一螺鏇形骨管,繞蝸軸捲曲兩周半。由蝸軸向管的中央伸出一片簿骨,叫骨質螺板。耳蝸外壁有螺鏇韌帶。骨質螺鏇板的游離緣連著一富有彈性的纖維膜,稱為基底膜,延伸到骨管對側壁與螺鏇韌帶相接。把耳蝸骨管分成上下兩部,上部稱前庭階,下部稱鼓階,兩管中充滿外淋巴液。前庭階的一端為前庭窗,鼓階一端為蝸窗。兩部分在蝸頂處的蝸孔相通。在骨質螺鏇板近底處有一薄膜,稱前庭膜,由前庭膜、基底膜和一部分螺鏇韌帶圍成膜質蝸管,管中充滿內淋巴液。螺鏇器(Corti氏器)是感受聲波刺激的聽覺感受器,由支持細胞和毛細胞等組成,毛細胞為聲波感受細胞,每個毛細胞均與神經纖維形成突觸聯繫。毛細胞的上方有鼓膜,與毛細胞的纖毛相接觸。外界聲波通過淋巴液而震動鼓膜,鼓膜又觸動了毛細胞,最後由毛細胞轉換成神經衝動經聽位神經而傳到聽覺中樞。
耳蝸微音器效應是把傳到耳蝸的機械振動轉變成聽神經纖維的神經衝動。在這一轉變過程中,耳蝸基底膜的振動是一個關鍵因素。它的振動使位於它上面的毛細胞受到刺激,引起耳蝸內發生各種過渡性的電變化,最後引起位於毛細胞底部的傳入神經纖維產生動作電位。
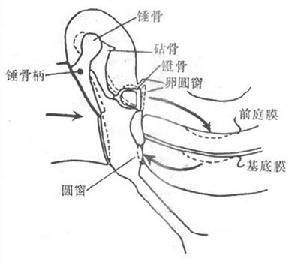
行波理論
 行波理論
行波理論當聲流振動通過聽骨鏈到達卵圓窗膜時,壓力變化立即傳給隔離蝸內液體和膜性結構;如果卵圓窗膜內移,前庭膜和基底膜也將下移,最後是鼓階的外淋巴壓迫圓窗膜外移;相反,當卵圓窗膜外移時,整個耳蝸內結構又作反方向的移動,於是形成振動。可以看出,在正常氣傳導的過程中,圓窗膜實際起著緩衝耳蝸內壓力變化的作用,是耳蝸內結構發生振動的必要條件。有人用直接觀察的方法,詳細記錄了聲音刺激引起的基底膜振動的情況,這對於了解基底膜振動的形式,以及這種振動在耳蝸接受不同頻率的聲音刺激時有何差異,提供了可靠的依據。觀察表明,基底膜的振動是以行波(traveling wave)的方式進行的,即內淋巴的振動首先是靠近卵圓窗處引起基底膜的振動,此波動再以行波的形式沿基底膜向耳蝸的頂部方向傳播,就像人在抖動一條綢帶時,有行波沿綢帶向遠端傳播一樣。下一步還證明,不同頻率的聲音引起的行波都從基底膜的底部,即靠近卵圓窗膜處開始,但頻率不同時,行波傳播的遠近和最大行波的出現部位有氣溫同,這就是振動頻率愈低,行波傳播愈遠,最大行波振幅出現的部位愈靠近基底膜頂部,而且在行波最大振幅出現後,行波很快消失,不再傳播;相反地,高頻率聲音引起的基底膜振動,只局限於卵圓窗附近。
不同頻率的振動引起的基底膜不同形式的行波傳播,主要由基底膜的某些物理性質決定的。基底膜的長度在人約為30mm,較耳蝸略短,但寬度在靠近卵圓窗處只有0.04mm,以且逐漸加寬;與此相對應,基底膜上的蚴鏇器的高度和重量,也隨著基底膜的加寬而變大。這些因素決定了基底膜愈靠近底部,共振頻率愈高,愈靠近頂部,共振頻率愈低;這就使得低頻振動引起的行波在向頂部傳播時阻力較小,而高頻振動引起的行波只限局在底部附近。
不同頻率的聲音引起的不同形式的基底膜的振動,被認為是耳蝸能區分不同聲音頻率的基礎。破壞動物不同部位基底膜的實驗和臨床上不同性質耳聾原因的研究,都證明了這一結論,亦即耳蝸底部受時主要影響高頻聽力,耳蝸頂部受損時主要影響低頻聽力。不能理解,既然每一種振動頻率在基底膜上都有一個特定的行波傳播範圍和最大振幅區,與這些區域有關的毛細胞和聽神經纖維就會受到最大的刺激,這樣,來自基底膜不同區域的聽神經纖維的神經衝動及其組合形式,傳到聽覺中樞的不同部位,就可能引起不同音調的感覺。
基底膜的振動怎樣使毛細胞受到刺激,毛細胞頂端的聽毛有些埋在蓋膜的膠狀物中,有些是和蓋膜的下面接觸;因蓋膜和基底膜的振動軸不一致,於是兩膜之間有一個橫向的交錯移動,使聽毛受到一個切向力的作用而彎曲。毛細胞聽纖毛的彎曲,是耳蝸中由機械能轉為電變化的第一步。
生物現象
 耳蝸效應是聽覺的核心
耳蝸效應是聽覺的核心當耳蝸接受聲音刺激時,在耳蝸及其附近結構又可記錄到一種特殊的電波動,稱為微音器電位。這是一種交流性質的電變化,在一定的刺激強度範圍內,它的頻率和幅度與聲波振動完全一致;這一現象正如向一個電話機的受話器或微音器(即麥克風)發聲時,它們可將聲音振動轉變為波形類似的音頻電信號一樣,這正是把耳蝸的這種電變化稱為微音器電位的原因。事實上,如果對著一個實驗動物和耳廓講話,同時在耳蝸引導它的微音器電位,並將此電位經放大後連線到一個揚聲器,那么揚聲器發出的聲音正好是講話的聲音!這一實驗生動地說明,耳蝸在這裡起著類似微音器的作用,能把聲波變成相應的音頻電信號。微音器電位的其它一些特點是:潛伏期極短,小於0.1ms;沒有不應期;對缺O2和深麻醉相對地不敏感,以及它在聽神經纖維變性時仍能出現等。
用微電極進入毛細胞的細胞內電變化記錄的實驗證明,所謂微音器電位就是多個毛細胞在接受聲音刺激時產生的感受器電位的複合表現;在記錄單一毛細胞跨膜電位的情況下,發現聽毛只要有0.1。的角位移,就可引起毛細胞出現感受器電位,而且電位變化的方向與聽毛受力的方向有關,亦即此電位既可是去極化的;這就說明了為什麼微音器電位的波動同聲波振動的頻率和幅度相一致。
由於聽毛的角位移和產生感受器電位之間只有一極短潛伏期,因而認為後者的產生是由於毛細胞頂部膜中有機械門控通道的存在,聽毛受力引起該處膜的輕微變形,就足以改變這種通道蛋白質的功能狀態,引起跨膜離子移動和相應的電位反應。在毛細胞,它的感受器電位可引起細胞基底部的遞質(可能是谷氨酸和門冬氨酸)釋放量的改變,進而引起分布在附近的耳蝸傳入纖維產生動作電位,傳向聽覺高級樞,產生聽覺。
至於內毛細胞和外毛細胞在功能上有何不同,有人首先注意到它們所接受的傳入纖維的數目有極大差異。人一側耳蝸內毛細胞的總數約為3500個,外毛細胞則有約15000個,但來自螺鏇神經節的約32000條聽神經傳入纖維中約有90%分布到內毛細胞的底部,這說明一個內毛細胞可接受多條傳入纖維的分布,而多個外毛細胞才能接受一個傳入纖維的軸突分支。因此一般認為,內毛細胞的作用是把不同頻率的聲音振動轉變為大量分布在它們底部的傳入纖維的神經衝動,向中樞傳送聽覺信息,而息細胞的作用近年來卻發現有些特殊。有人發現毛細胞在基底膜振動和聽毛受力而出現微音器電位時,此細胞可產生形體長短的快速改變,超極化引起細胞伸長,去極化引起細胞縮短,它們的形體改變因此也和外來聲音振動的頻率和振幅同步。據認為,外毛細胞的這種形體改變可以使所在基底膜部分原有的振動增強,亦卻對行經該處的行波起放大作用,這顯然使位於該部分基底膜上的內毛細胞更易受到刺激,提高了對該振動頻率的敏感性。外毛細胞因膜內外電位差改變引起的機制尚不清楚,但這使得基底膜不僅僅是以固定的結構“被動”地對外界的振動產生行波,它還可以“主動”地增強行波的振動幅度。
