
有髓
 神經纖維
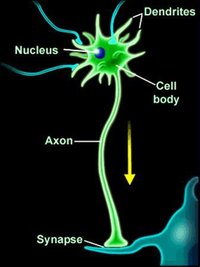
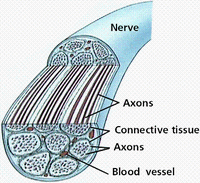
神經纖維有髓神經纖維的軸突外包有一層髓鞘,髓鞘外有一層神經膜。髓鞘與神經膜是一個神經膜細胞的兩個部分:神經膜是細胞含有細胞質和細胞核的部分,髓鞘是神經膜細胞的細胞膜反覆纏繞在軸突周圍形成的多層膜結構。有髓神經纖維每隔一定的距離,髓鞘便有間斷,此處變窄,稱神經纖維節或郎飛氏結。兩個郎飛氏結之間的一段,稱結間段。神經纖維愈粗,結間段愈長。一個神經膜細胞構成一個結間段的髓鞘和神經膜。
髓鞘由蛋白質和脂類組成,在H-E染色的切片中,髓鞘的脂類物被酒精溶解,只留下網狀的蛋白結構。用鋨酸處理後,髓鞘被染成黑色。在神經纖維縱切面上,可見髓鞘內有斜行或漏斗狀裂隙,稱施蘭氏裂或髓鞘切跡,它是神經膜細胞質通入髓鞘內的通道,有利於髓鞘和軸突的物質更新。
若把纏繞在軸突上的神經膜細胞展開,細胞的形態如同一梯形的扁囊,梯形的短底靠近軸突。扁囊的大部分,兩層細胞膜緊貼,但在囊的四周和中間具有細胞質的通道。梯形長底的細胞質通道稱外環,此處含有細胞核及各種細胞器,是構成神經膜的部分。短底的通道稱內環,緊纏軸突。兩側邊的通道稱節旁環,纏在神經纖維節的兩旁。纏有節旁環的軸突部分稱節旁部。在神經膜細胞中和內環、外環間相連的細胞質通道纏繞在軸突上,構成髓鞘切跡。髓鞘切跡使細胞質能與構成髓鞘的各部分細胞膜接觸,從而參與髓鞘結構的新陳代謝。 構成髓鞘的細胞膜沒有鈉泵和離子通道,這樣使有髓鞘包裹的軸突部分。離子不能通過。有髓神經纖維神經衝動的傳播是從一個神經纖維節跳到相鄰的另一個神經纖維節,呈跳躍式傳導。節間段愈長,跳躍的距離也就愈遠傳導速度也就愈快。因此,軸突有了髓鞘可大大增快神經衝動的傳播速度,這就是髓鞘所起的作用。
傳導功能
 神經纖維
神經纖維神經纖維的功能是傳導興奮或神經衝動,其傳入纖維將感受器的興奮傳到中樞,而傳出纖維又將中樞的興奮傳至效應器。
傳導特點
神經纖維在結構上及生理功能上應該是完整的,即使結構完整,而局部環境發生變化,如麻醉、低溫,也可阻滯衝動的傳導。
每條神經乾包含的任何一條神經纖維都沿本身傳導衝動,與相鄰纖維相互隔絕,不相干擾,這種絕緣性使神經調節更為精確。
神經纖維任何一點受到的刺激所產生的衝動可沿纖維向兩端即雙向傳導。
神經纖維有相對不疲勞性,始終保持傳導能力。
傳導機制
有髓神經纖維的髓鞘在郎飛結處缺失,當某一郎飛結興奮時,這一區域就出現除極。髓鞘主要由脂類物質構成,具有較大的阻抗。局部電流只能沿軸突流動,直到下一個未興奮的郎飛結處才穿出,然後沿髓鞘外回到原先興奮的部位,這樣在已興奮的郎飛結與鄰近安靜的郎飛結之間形成局部電流,可以興奮下一個未興奮的郎飛結,這就是所謂的跳躍傳導。
無髓神經纖維的傳導速度比有髓纖維要慢。神經纖維某一段受刺激而興奮,立即出現峰電位,該處膜電位暫時倒轉而除極,呈內正外負,但鄰近部位仍處於安靜時的極化狀態,呈內負外正,因此興奮部位與鄰近安靜部位之間形成電位差,導致電荷移動,產生局部電流,使鄰近安靜部位興奮,峰電位沿整個神經纖維傳導。神經纖維直徑越大,纖維內縱向阻抗越小,局部電流增大,而傳導速度加快。
軸漿
 神經纖維
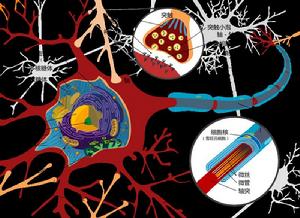
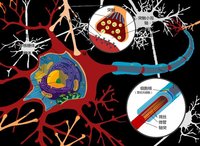
神經纖維神經元的細胞體與軸突是一個整體,胞體和軸突之間必須經常進行物質運輸和交換。實驗證明,軸突內的軸漿是經常在流動的。軸漿流動是雙向的,一方面部分軸漿由胞體流向軸突末梢,另一方面部分軸漿由軸突末梢反向流向胞體。胞體內具有高速度合成蛋白質的結構,其合成的物質借軸漿流動向軸突末梢運輸;而反向的軸漿流動可能起著反饋控制胞體合成蛋白質的作用。在組織培養或在體的神經纖維中,用顯微鏡觀察確實見到軸漿內顆粒具有雙向流動的現象。用同位素標記的胺基酸注射到蛛網膜下腔中,可以見到注射物質首先被神經元的細胞體報到,而在胞體內出現,然後逐漸在軸突近端軸漿內出現,最後在遠端軸漿內出現,說明軸漿在流動。如果軸突中斷,思漿雙向流動被阻斷,則遠側斷端和近側斷端及胞體都受到影響;因此變性反應不僅發生在遠端正,也發生在胞體。 自胞體向軸突末梢的軸漿運輸分兩類。一類是快速軸漿運輸,指的是具有膜的細胞器(線粒體、遞質囊泡、分泌顆粒等)的運輸,在猴、貓等動物的坐骨神經內其運輸速度為410mm/d。另一類是慢速軸漿運輸,指的是由胞體合成的蛋白質所構成的微管和微絲等結構不斷向前延伸,其他軸漿的可溶性成分也隨之向前運輸,其速度為1-12mm/d。
軸漿流動的機制目前還不十分清楚。在缺氧、氰化物毒化等情況下,神經纖維的有氧代謝擾亂使ATP減少到50%以下時,快速軸獎學金流動即停止,說明它是一種耗能過程。有人提出與肌肉收縮滑行理論相似的假說,來解釋快速軸漿流動。認為囊泡等有膜的細胞器的運輸與微管成微絲的功能有關,微管的成分與肌纖蛋白相似,微管上含有結合點和ATP,囊泡膜上有ATP酶和能與微管相附著的結合點;ATP酶作用於ATP,後者放出能量使微管與囊泡膜發生附著結合,而後又脫離接觸,如此推動囊泡不斷與下一個結合點相附著,造成囊泡等有膜細胞器沿著微管向前推移。
由軸突末梢向細胞體方向的逆向軸漿流動了解得比較少。這種逆向流動的速度約為快速順向運輸速度的一半左右。有人認為,破傷風毒素、狂犬病病毒由外周向中樞神經系統轉運的機制,可能就是逆向軸漿流動。運用辣根過氧化酶方法研究神經纖維的發源部位,其原理也是因為辣根過氧化酶能被軸突末梢攝取,並由軸漿流動轉運到神經纖維的細胞體。
潰變
 神經纖維
神經纖維神經纖維受損傷如神經被切斷後,切斷處遠側段的神經纖維全長發生潰變(degeneration),軸突和髓鞘碎裂和溶解。與胞體相連的近側段則發生逆行性潰變,即軸突和髓鞘的斷裂溶解由切斷處向胞體方向進行,潰變一般只發展到鄰近斷端的第一側支終止。神經元的胞體腫脹,細胞核從中央移到胞體邊緣,胞質內尼氏體明顯減少,故胞質著色淺淡。
再生
 神經纖維
神經纖維傷神經元胞體是細胞的營養中心,只有在胞體沒有死亡的條件下才有纖維再生的可能。胞體約於損後第3周開始恢復,胞質內的尼氏體重新出現,胞體腫脹消失,胞核恢復中央位置。胞體的完全恢復約需3~6個月,恢復中的胞體不斷合成新的蛋白質及其他產物輸向軸突,使殘留的近側段軸突末端生長出許多新生的軸突支芽。
1、周圍神經纖維的再生切斷處遠側段的周圍神經纖維,雖然其軸突和髓鞘發生潰變,但包裹神經纖維的基膜仍保留呈管狀。此時施萬細胞大量增生,一面吞噬解體的軸突和髓鞘,一面在基膜管內排列成細胞索,靠近斷口處的施萬細胞還形成細胞橋把兩斷端連線起來。從近側段神經纖維軸突末端長出的軸突支芽,越過此施萬細胞橋,進入基膜管內,當其中一支沿著施萬細胞索生長併到達原來神經纖維末梢所在處,則再生成功。施萬細胞和基膜對軸突的再生起重要的誘導作用。
2、中樞神經纖維的再生中樞神經纖維的再生比周圍神經困難。神經纖維無施萬細胞,亦無基膜包裹。當中樞神經纖維受損傷時,星形膠質細胞增生肥大,在損傷區形成緻密的膠質瘢痕,大多數再生軸突支不能越過此膠質瘢痕;即使能越過,也沒有如同周圍神經纖維那樣的基膜管和施萬細胞索引導再生軸突到達目的地。所以,中樞神經纖維的損傷常導致脊髓或腦功能的永久性喪失。不少科學家為研究神經再生進行不懈的努力,已注意到一類能促進神經生長的化學物質稱神經營養因子的作用。同時又根據胚胎神經元容易生長的化學物質稱神經營養因子的作用。同時又根據胚胎神經元容易生長及周圍神經能再生的特點,把胚胎腦組織、周圍神經或周圍神經的組分(如基膜或基膜的化學成分)移植到腦內,以期促進中樞神經再生。
