 抑制基因
抑制基因Bcl-2基因
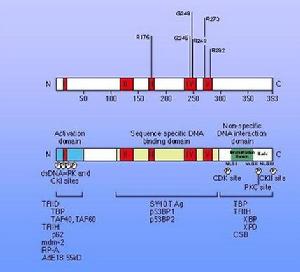
在細胞凋亡的相關調控基因的研究方面,Bcl-2基因是目前研究的最深入、最廣泛的凋亡調控基因之一。Bcl-2基因最初是在非霍奇金濾泡狀B細胞淋巴瘤中分離出來的,它是在14號與18號染色體易位的斷點上被發現的。Bcl-2基因編碼一個25-26KD的蛋白,其C端的21個疏水胺基酸組成一個延伸的鏈狀結構。這個鏈可以插到細胞的膜結構中,這一結構特點與Bcl-2調節細胞凋亡的方式和能力非常有關。已經證實Bcl-2存在於線粒體外膜、核膜和內質網膜上。根據對Bcl-2家族成員蛋白的結構研究,發現其共同點是在蛋白鏈上都存在三個結構基序,分別稱為BH1、BH2和BH3結構域。和Ced-9一樣,Bcl-2基因屬於一類新的癌基因家族成員,通過有效抑制許多不同類型細胞中許多不同類型的凋亡刺激誘導的細胞凋亡,延長細胞活力而發揮其生物學作用,對細胞周期的進程不發生影響。這說明它在細胞凋亡調控機制中起著十分重要的作用,可能是許多因子作用的共同分子基礎。但是Bcl-2基因如何抑制細胞凋亡的機制尚不清楚。最近發現Bcl-2可與細胞中的bax蛋白質結合構成不同的二聚體(Bcl-2/ Bcl-2、Bcl-2/bax、bax/bax),通過它們之間的不同比例來調節細胞凋亡。用EB病毒來感染細胞後可使細胞發生永生或生存期延長,這是因為EB病毒促使Bcl-2表達的結果。套用原位雜交和免疫組化方法對鼻咽癌組織和體外培養的細胞進行研究發現,鼻咽癌組織及其體外培養細胞出現bcl-2的過表達。
Bcl-2基因家族是目前廣泛研究的一類細胞凋亡相關基因,其表達和調控是影響細胞凋亡的關鍵因素之一,在細胞凋亡信號轉導途徑中發揮質量要作用。通過上調促凋亡的Bcl-2的基因表達,從而在轉錄水平調節凋亡相關蛋白的表達以影響凋亡的發生。Bcl-2家族成員都含有1-4個Bcl-2同源結構域(BH1-4),並且通常有一個羧端跨膜結構域。其中BH4是抗凋亡蛋白所特有的結構域,BH3是與促進凋亡有關的結構域。根據功能和結構可將Bcl-2基因家族分為兩類,一類是抗凋亡的(anti-apoptotic),如:Bcl-2、Bcl-xl、Bcl-w、Mcl-1;一類是促進凋亡的(Pro-Apoptotic),如:Bax、Bak、Bad、Bid、Bim,在促凋亡蛋白中還有一類僅含BH3結構,如Bid、Bad。目前已經在哺乳動物、線蟲和細菌中發現並鑑定出了bcl-2家族的20餘種蛋白質,根據它們在細胞凋亡中的作用可分為兩類:一類是抗凋亡蛋白,包括Bcl-2、Bcl-xL、Bcl-W、Mcl-l、AI、Boo和Ced-9等十餘個成員;另一類是促凋亡蛋白,包括Bax、Bak、Bcl-xS、Bid、Bad和Egl-l等十餘個成員。Bcl-2是迄今研究得最深入、最廣泛的凋亡調控基因之一,編碼產物Bcl-2蛋白可抑制細胞凋亡,參與細胞增殖與凋亡動態平衡的調控。
突變基因的表型效應由於第二個突變基因的出現而恢復正常時,稱後一突變基因為前者的抑制基因。回復突變使突變基因的脫氧核糖核酸(DNA)分子結構恢復正常;抑制基因則並不改變突變基因的DNA分子結構,而只是使突變型的表型恢復正常。抑制基因一般用符號Su代表。例如用Hw代表黑腹果蠅的毛翅(Hairywing,Hw)突變型,則Su-Hw代表Hw的一個抑制基因。當抑制作用發生在同一基因中時,這種抑制作用稱為基因內抑制,屬於不同基因的抑制作用則稱基因間抑制。
基因抑制分類
 抑制基因
抑制基因針對移碼突變的基因內抑制大腸桿菌噬菌體T4經吖啶類染料處理後可以得到一對核苷酸減少(或增加)的rⅡ基因的移碼突變型。在這一對核苷酸不改變的情況下,附近位置上另一對核苷酸的增加(或減少)能使表型恢復正常(見遺傳密碼),這便是針對移碼突變的基因內抑制,抑制基因本身也常是移碼突變型。針對無義突變的基因內抑制密碼子AAG代表賴氨酸,由於置換突變而成為無義密碼子UAG時,翻譯便到此停止而帶來突變型表型。在第一對核苷酸不改變的情況下,由於第三對核苷酸的改變而使密碼子成為UAC時,便在這位置上出現酪氨酸並使翻譯正常進行。只要由賴氨酸改變為酪氨酸不影響這一基因產物的活性,表型便得以恢復。這是針對無義突變的基因內抑制。
基因間抑制通過代謝補償的基因間抑制假定某一基因突變使一種蛋白質變為容易為另一物質所抑制,因而使野生型表型不能出現,再假定另一基因發生突變後這抑制物不再產生,那么後一突變基因便是前一突變基因的抑制基因。假定某一突變基因使某一代謝產物不能形成,再假定另一突變基因使同一代謝產物由另一途徑合成,那么它也是前一突變基因的抑制基因。如果某一代謝中間產物具有某種表型效應,假定第一個突變基因中斷了這一中間產物以後的代謝途徑而使中間產物積累,再假定另一突變基因使中間產物出現以前的代謝途徑中斷,那么它也成為前一突變基因的抑制基因。例如粗糙脈孢菌的腺嘌呤缺陷型(adenineless,ade)35203產生一種由腺核苷酸合成的代謝中間產物轉變來的紫色色素(見圖)。在這一基因不發生回復突變的情況下,另外三個腺嘌呤缺陷型27663、28610、44411中的任何一個都抑制紫色色素的產生而恢復了野生型的不產色素這一性狀。對於突變型35203的產生紫色色素這一性狀來講,突變基因27663、28610、44411都是它的抑制基因。
通過翻譯校正的基因間抑制某些由置換突變產生的無義突變型,可以由於相應的突變型轉運核糖核酸(tRNA)的校正作用而恢復正常的表型。例如酪氨酸密碼子是UAC,酪氨酸tRNA反密碼子是AUG。置換突變使UAC變為無義密碼子UAG後翻譯便到此停止。如果酪氨酸tRNA基因發生突變而使它的反密碼子由AUG變為AUC,這一反密碼子便能識別無義密碼子UAG,可是它的3′端上仍然攜帶著酪氨酸。因此這一突變型tRNA能使無義突變密碼子位置上照常出現酪氨酸而使翻譯正常進行。這裡酪氨酸tRNA的突變基因便是前一無義突變型的抑制基因。
任何一個突變型只要它和野生型只有一對核苷酸的差別,都可以由相應的tRNA基因突變型的校正作用而恢復野生型表型。抑制基因的專一性和表型效應各種抑制基因的作用可以有不同的專一性。果蠅的毛翅抑制基因Su-Hw除了抑制毛翅以外,對於分叉剛毛(bifid,bi),翅脈中斷(cubitusinterruptus,ciD)也有一定程度的抑制作用;星眼抑制基因則只對星眼突變(star,S)有抑制作用。脈孢菌的紫色腺嘌呤缺陷型的抑制基因只抑制該突變型的紫色表型而不抑制對於腺嘌呤的需要這一表型。琥珀突變型抑制基因的抑制作用針對無義密碼子UAG,而不論這一無義突變發生在哪一基因中,所以它又稱為超抑制基因。相反地,許多抑制基因的作用是座位專一的,它能抑制某一突變基因的表型,不論突變發生在這基因的哪一位點。某些抑制基因的作用則是位點專一的,它對被抑制基因的抑制作用只限於某些位點上的突變。例如大腸桿菌的突變型dnaAts是在高溫中不能複製DNA的突變型。rpoB是它的抑制基因,每一個rpoB突變只抑制某一些位點發生突變的dnaAts突變型,而不抑制同一基因的某些其他位點的突變型。一般認為位點專一性是由於抑制基因與被抑制基因所編碼的兩種蛋白質分子的相互作用只限於某些相對應的部位的緣故。某些抑制基因能夠抑制許多突變基因的表型,例如琥珀突變型抑制基因。相反地,一個基因的表型也可以被幾個不同的抑制基因所抑制,例如脈孢菌紫色腺嘌呤缺陷型至少有三個抑制基因。
某些抑制基因本身並沒有表型效應,例如果蠅的毛翅抑制基因。某些抑制基因本身具有突變型表型效應,例如大腸桿菌的色氨酸合成酶A亞基基因的突變型A446對於另一個A亞基突變型A46是抑制基因,A46對A446來講也是抑制基因。脈孢菌的紫色腺嘌呤缺陷型的幾個抑制基因本身也都是腺嘌呤缺陷型。大腸桿菌中由於tRNA基因發生突變的抑制基因則雖然沒有一般的突變型表型,可是普遍地降低生活力。
抑制基因或是隱性的,或是顯性的。在細菌等單倍體生物中,不論抑制基因是隱性或顯性,往往可以從被它抑制的突變型──假的回覆體中發現。這些假的回覆體和野生型雜交的子代中總是或多或少地出現原來的突變型;相反地,真的回覆體和野生型雜交的子代中則不出現原來的突變型。在二倍體的高等生物中,按照子二代中出現野生型和突變型的比數便可以判斷抑制基因的存在以及抑制基因的顯隱性關係。
