
病因介紹
 脊髓灰質炎病毒
脊髓灰質炎病毒髓灰質炎病毒經口進入人體後,即侵入咽部和腸道的淋巴組織,則形成隱性感染;抗體低下病毒則進入血循環,引起病毒血症。若機體缺乏免疫力,病毒隨血流經血腦屏障侵入中樞神經系統,並沿神經纖維擴散,引起無癱瘓期症狀;如果運動神經元受損嚴重,則導致肌肉癱瘓,引起癱瘓期症狀。
病原菌脊髓灰質炎病毒屬於微小核糖核酸(RNA)病毒科(picornaviridae)的腸道病毒屬(enterovirus)。此類病毒具有某些相同的理化生物特徵,在電鏡下呈球形顆粒相對較小,直徑20~30nm,呈立體對稱12面體。病毒顆粒中心為單股正鏈核糖核酸,外圍32個衣殼微粒,形成外層衣殼,此種病毒核衣殼體裸露無囊膜。核衣殼含4種結構蛋白VP1、VP3和由VP0分裂而成的VP2和VP4.VP1為主要的外露蛋白至少含2個表位(epitope),可誘導中和抗體的產生,VP1對人體細胞膜上受體(可能位於染色體19上)有特殊親和力,與病毒的致病性和毒性有關。VP0最終分裂為VP2與VP4,為內在蛋白與RNA密切結合,VP2與VP3半暴露具抗原性。
人類腸道病毒的分子結構大致相同。脊髓灰質炎的分子生物學研究較深入,對腸道病毒基因組序列化發現約含7450個核苷酸,分為三個區域:5'未端有743個核苷酸,下接一個編碼區約6625個核苷酸,最後為3'poly(A)端區,長短不一,而與RNA感染性有關。5'端與小病毒編碼蛋白(VPg)相接,與啟動RNA合成有關。
由於Polio病毒無囊膜,外衣不含類脂質,故可抵抗乙醚、乙醇和膽鹽。在pH3.0~10.0病毒可保持穩定,對胃液、腸液具有抵抗力,利於病毒在腸道生長繁殖。病毒在人體外生活力很強,污水及糞便中可存活4~6月,低溫下可長期存活,-20℃~-70℃可存活數年,但對高溫及乾燥甚敏感,煮沸立即死亡,加溫56℃半小時滅活,紫外線可在0.5~1小時內將其殺死。各種氧化劑(漂白粉、過氧化氫、氯胺、過錳酸鉀等),2%碘酊、甲醛、升汞等都有消毒作用;在含有0.3~0.5PPM游離氯的水中10分鐘滅活,1∶1000高錳酸鉀及2%碘酊,3%~5%甲醛均可很快使病毒滅活,丙酮、石炭酸的滅活作用較緩慢。70%酒精、5%來蘇爾無消毒作用,抗生素及化學藥物也無效。
目前認為人是脊髓灰質炎病毒的唯一天然宿主,因人細胞膜表面有一種受體對病毒VPI具有特異親和力。實驗感染也只發現猩猩和猴對其易感。某些毒株可使乳鼠發病。越低級的靈長類動物越不易發生神經系統感染,而易得腸道感染。組織培養以人胚腎、人胚肺、人羊膜及猴腎細胞最為敏感,在Hela細胞中也易培養,Hela細胞膜上有對三個型Polio病毒的共同受體,病毒引起細胞圓縮、脫落等病變。
脊髓灰質炎病毒按抗原性不同可分為1、2、3型,型間偶有交叉免疫。不同病毒株的性能可略有差異,如有的毒株具有對神經組織的親和力,可引起麻痹,不同毒株之間這種親和力可相差1000倍之巨,而同型減毒活疫苗株則幾乎已無此毒性,但對熱的敏感性則增加了,並可出現細微的抗原性差異,且經人腸道反覆傳代數月至數年後疫苗株可產生變異如使人體排病毒時間延長,對神經細胞毒性增強,使人體產生較多干擾素,故在鑑別野毒株或疫苗株時應以分子生物學技術的結果為準。每型病毒含有二種特異抗原:一為存在於成熟病毒體內的D(dense)抗原,另一種為與缺乏RNA的空殼病毒顆粒有關的C(coreless)抗原,存在於前衣殼(procapsid)中,病毒在機體中和抗體作用下D抗原性可轉變為C抗原性,並失去使易感細胞發生感染的能力。
病毒型別
已知脊髓灰質炎病毒有三個血清型,這三型病毒的核苷酸序列已經清楚,總的核苷酸數目為7500個左右。雖然有71%左右的核苷酸為三型脊髓灰質炎病毒所共有,但不相同的核苷酸序列卻都位於編碼區內,因此三型病毒間中和試驗無交叉反應。
抗原性狀
 脊髓灰質炎病毒
脊髓灰質炎病毒用補體結合試驗可查出病毒有兩種抗原,一種稱為D(緻密)抗原,另一種稱為
C(無核心)抗原。前者存在於成熟的、有感染性的病毒顆粒中,是該病毒的中和抗原,具有型特異性。C抗原存在於經過56℃滅活,或者未成熟的空心病毒顆粒中,是一種耐熱的抗原成份,與三型病毒的抗血清均呈補體結合陽性反應。
流行病學
 脊髓灰質炎病毒
脊髓灰質炎病毒溫帶多見脊髓灰質炎,終年散發,以夏秋為多,可呈小流行或釀成大流行,熱帶則四季發病率相似。世界各國都有發病,但在普種疫苗地區發病率大大減少,幾乎無發病(如北歐芬蘭、瑞士、荷蘭等國),中國1976~1980年平均發病率也已降至0.7/10萬;尤以大中城市嬰幼兒服疫苗率已達80%以上地區發病率下降為快,如江蘇省已從1956年的10.51/10萬降至1982年的0.2/10萬。未用疫苗地區
則仍有流行。以往以1型為多,而近2、3型病毒相應多見。流行時以無症狀的隱性感染及不發生癱瘓的輕症較多。在熱帶、人口密聚及未廣泛服用疫苗地區,仍以1~5歲小兒發病率最高。自嬰幼兒廣泛採用疫苗後,世界各地發病年齡有逐步提高趨勢,以學齡兒童和少年為多,成人患者也有所增加。1歲以內發病者也增多。傳染源為病人及無症狀的帶病毒者,後者不僅人數眾多,又不易被發現和控制,因而對本病的散布和流行起著重要作用。在兒童中癱瘓病例與隱性感染及無癱瘓病例之比可高達1∶1000,成人中也可達1∶75。流行時幼托機構中感染率可高達100%。早在發病前3~5日患者鼻咽分泌物及糞便內已可排出病毒。咽部主要在病初1周內排出病毒,故通過飛沫傳播的時間亦短,而糞便中排出病毒不僅時間早(病前10天)、量多、且可持續2~6周,甚至長達3~4個月,因此糞便污染飲食,經口攝入為本病主要傳播途徑。直接或間接污染病毒的雙手、用品、玩具、衣服及蒼蠅等皆可成為傳播媒介,飲水污染常引起爆發流行。
 脊髓灰質炎病毒
脊髓灰質炎病毒感染後人體對同型病毒能產生較持久的免疫力,血清中最早出現特異型IgM,
兩周后出現IgG(中和抗體)。唾液及腸道產生分泌型IgA。中和抗體水平在起病後2~3周到達高峰,1~2年內漸下降,但一直保持一定水平,不僅可保護患者免遭同型病毒感染,對異型病毒也具低保護力。此外,此病毒具有C和D兩種抗原。C抗體病後出現早而在病程1~2周后即下降,D抗體出現較遲,2個月達高峰,保持2年左右,有型特異性。特異抗體可通過胎盤(IgG)及母乳(含分泌型IgA)自母體傳新生兒,此種被動免疫在出生後6個月中漸漸消失。年長兒大多經過隱性感染獲得自動免疫力,抗體水平再度增長;到成人時大多數已具有一定免疫力。
病毒分析
為及時發現和阻斷疫苗衍生脊髓灰質炎(脊灰)病毒(vaccine-derived polioviruses, VDPVs)循環、脊灰野病毒的輸入和能夠引起臨床脊灰的其它脊灰疫苗相關病毒,做好貴州省無脊灰狀態下脊灰病毒學監測工作,採用病毒分離、鑑定與核苷酸序列測定和分析方法。
對貴州省貞豐縣及周圍10個縣2003~2004年報告的急性弛緩性麻痹(AFP)病例及2004年接觸者糞便標本的病毒學監測的結果進行了分析。對收集到的105例AFP病例和47例密切接觸者的278份便標本進行了病毒學監測,結果從66例中共分離到腸道病毒(EV)66株,陽性率為43.4%,其中脊灰病毒(PV)29例,分離率為19.1%,非脊灰腸道病毒(NPEV)37例,分離率為24.3%.29例PV經中國疾病預防控制中心病毒病預防控制所國家脊灰實驗室鑑定,24例為疫苗相似株,5例為脊灰Ⅰ型VDPVs,這5例均為2004年從貞豐縣所分離到。
貞豐縣及周圍縣EV陽性檢出率(43.4%)高於2003~2004年全省水平(22.9%~24.6%),2004年PV分離率比2003年高達2.6倍,29株PV中單個Ⅰ型占34.5%,明顯高於往年(2000~2002年全省平均4.1%)。本研究提示,Ⅰ型VDPVs在貞豐縣引起了循環(cVDPVs),通過口服脊灰疫苗強化免疫已經阻斷cVDPVs的傳播。人群中PV和NPEV帶毒率明顯增高,非VDPVs引起的臨床符合脊灰病例不容忽視,應加強脊灰病毒學監測數據的分析和早期疫情預警工作。
發病機制
 脊髓灰質炎病毒發病機制
脊髓灰質炎病毒發病機制一、病原菌脊髓灰質炎病毒屬於微小核糖核酸(RNA)病毒科(picornaviridae)的腸道病毒屬(enterovirus)。此類病毒具有某些相同的理化生物特徵,在電鏡下呈球形顆粒相對較小,直徑20~30nm,呈立體對稱12面體。病毒顆粒中心為單股正鏈核糖核酸,外圍32個衣殼微粒,形成外層衣殼,此種病毒核衣殼體裸露無囊膜。核衣殼含4種結構蛋白VP1、VP3和由VP0分裂而成的VP2和VP4.VP1為主要的外露蛋白至少含2個表位(epitope),可誘導中和抗體的產生,VP1對人體細胞膜上受體(可能位於染色體19上)有特殊親和力,與病毒的致病性和毒性有關。VP0最終分裂為VP2與VP4,為內在蛋白與RNA密切結合,VP2與VP3半暴露具抗原性。
人類腸道病毒的分子結構大致相同。脊髓灰質炎的分子生物學研究較深入,對腸道病毒基因組序列化發現約含7450個核苷酸,分為三個區域:5'未端有743個核苷酸,下接一個編碼區約6625個核苷酸,最後為3'poly(A)端區,長短不一,而與RNA感染性有關。5'端與小病毒編碼蛋白(VPg)相接,與啟動RNA合成有關。
由於Polio病毒無囊膜,外衣不含類脂質,故可抵抗乙醚、乙醇和膽鹽。在pH3.0~10.0病毒可保持穩定,對胃液、腸液具有抵抗力,利於病毒在腸道生長繁殖。病毒在人體外生活力很強,污水及糞便中可存活4~6月,低溫下可長期存活,-20℃~-70℃可存活數年,但對高溫及乾燥甚敏感,煮沸立即死亡,加溫56℃半小時滅活,紫外線可在0.5~1小時內將其殺死。各種氧化劑(漂白粉、過氧化氫、氯胺、過錳酸鉀等),2%碘酊、甲醛、升汞等都有消毒作用;在含有0.3~0.5PPM游離氯的水中10分鐘滅活,1∶1000高錳酸鉀及2%碘酊,3%~5%甲醛均可很快使病毒滅活,丙酮、石炭酸的滅活作用較緩慢。70%酒精、5%來蘇爾無消毒作用,抗生素及化學藥物也無效。
目前認為人是脊髓灰質炎病毒的唯一天然宿主,因人細胞膜表面有一種受體對病毒VPI具有特異親和力。實驗感染也只發現猩猩和猴對其易感。某些毒株可使乳鼠發病。越低級的靈長類動物越不易發生神經系統感染,而易得腸道感染。組織培養以人胚腎、人胚肺、人羊膜及猴腎細胞最為敏感,在Hela細胞中也易培養,Hela細胞膜上有對三個型Polio病毒的共同受體,病毒引起細胞圓縮、脫落等病變。
脊髓灰質炎病毒按抗原性不同可分為1、2、3型,型間偶有交叉免疫。不同病毒株的性能可略有差異,如有的毒株具有對神經組織的親和力,可引起麻痹,不同毒株之間這種親和力可相差1000倍之巨,而同型減毒活疫苗株則幾乎已無此毒性,但對熱的敏感性則增加了,並可出現細微的抗原性差異,且經人腸道反覆傳代數月至數年後疫苗株可產生變異如使人體排病毒時間延長,對神經細胞毒性增強,使人體產生較多干擾素,故在鑑別野毒株或疫苗株時應以分子生物學技術的結果為準。每型病毒含有二種特異抗原:一為存在於成熟病毒體內的D(dense)抗原,另一種為與缺乏RNA的空殼病毒顆粒有關的C(coreless)抗原,存在於前衣殼(procapsid)中,病毒在機體中和抗體作用下D抗原性可轉變為C抗原性,並失去使易感細胞發生感染的能力。
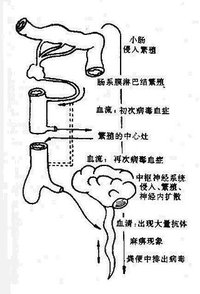
二、發病機理脊髓灰質炎病毒自口、咽或腸道黏膜侵入人體後,一天內即可到達局部淋巴組織,如扁桃體、咽壁淋巴組織、腸 壁集合淋巴組織等處生長繁殖,並向局部排出病毒。若此時人體產生多量特異抗體,可將病毒控制在局部,形成隱性感染;否則病毒進一步侵入血流(第一次病毒血症),在第3天到達各處非神經組織,如呼吸道、腸道、皮膚黏膜、心、腎、肝、胰、腎上腺等處繁殖,在全身淋巴組織中尤多,並於第4日至第7日再次大量進入血循環(第二次病毒血症),如果此時血循環中的特異抗體已足夠將病毒中和,則疾病發展至此為止,形成頓挫型脊髓灰質炎,僅有上呼吸道及腸道症狀,而不出現神經系統病變。少部分患者可因病毒毒力強或血中抗體不足以將其中和,病毒可隨血流經血腦屏障侵犯中樞神經系統,病變嚴重者可發生癱瘓。偶爾病毒也可沿外周神經傳播到中樞神經系統。特異中和抗體不易到達中樞神經系統和腸道,故腦脊液和糞便內病毒存留時間較長。因此,人體血循環中是否有特異抗體,其出現的時間早晚和數量是決定病毒能否侵犯中樞神經系統的重要因素。
多種因素可影響疾病的轉歸,如受涼、勞累、局部刺激、損傷、手術(如預防注射、扁桃體截除術、撥牙等),以及免疫力低下等,均有可能促使癱瘓的發生,孕婦如得病易發生癱瘓,年長兒和成人患者病情較重,發生癱瘓者多。兒童中男孩較女孩易患重症,多見癱瘓。
純化步驟
 純化步驟
純化步驟1、在大型裝置中培養細菌。為獲得足夠的用於化學研究的病毒,必須製備大量病毒。每種病毒的培養都有不同的步驟。脊髓灰質炎病毒是在人或靈長類動物細胞系中培養的,可以培養在一大瓶中沿壁生長的單層細胞上,也可以在一個輕輕攪拌的細胞懸液中。在有利病毒生長的條件下,每個宿主細胞能生產10~1000的感染單位的病毒,即每毫升一個感染單位的滴度(或稱效價)。
2、富含病毒培養液的去除。病毒的生長和釋放伴隨著細胞的破損。在病毒複製完成的時候,培養液中大多數細胞已經變成了細胞碎片。為使所有病毒分子完全釋放,要通過反覆冷凍—融化的循環過程使細胞進一步破碎。然後將富含病毒的液體從培養瓶轉移到離心試管中。低速離心除去大分子的細胞殘骸。
3、通過沉澱將病毒物質濃縮。由於病毒是蛋白質性質的,它們能通過蛋白質沉澱法沉澱。脊髓灰質炎病毒可通過高濃度的NH4Cl(每毫升培養液中加0.4g)沉澱下來。這一過程要在低溫下進行,NH4Cl要在攪拌下緩慢加到培養液中。由於病毒蛋白沉降,溶液將變得渾濁。將沉澱物通過低速離心(2000g,1-2hr)。然後將含有病毒分子的沉澱物在少量磷酸鹽緩衝液中再次懸浮培養。這一步驟可濃縮病毒大約10倍,99%以上的病毒粒子可沉澱回收。
4、通過密度梯度離心最終純化病毒。脊髓灰質炎病毒可以通過蔗糖或CSCl的密度梯度離心純化。在後者中,病毒在離心試管中成為一個分離帶,這一過程與對DNA的分離相同,離心須要高速,即120000g,且要進行好幾個小時,最好是一整夜。離心管中的液體通過試管底部的一點點滴出來,要檢測每一滴的病毒滴度。在合適的離心條件下,所有的病毒分子都集聚在一或兩個組分中,所以它們是非常純的。結晶脊髓灰質炎病毒。如果能在合適的條件下濃縮大量且高純度的病毒,就可以獲得病毒晶體。這種晶體為化學分析提供了很好的材料,且易於通過X射線衍射技術進行結構分析。
臨床表現
 脊髓灰質炎病毒
脊髓灰質炎病毒潛伏期一般5~14天(3~35天)。臨床症狀輕重不等,以輕者較多;多數可毫無症狀,倮可從鼻咽分泌物及大便中排出病毒,並可產生特異抗體9稱無症狀型或隱匿型,或隱性感染)。少數病人可出現弛緩性癱瘓,按癱瘓病人的病情發展過程,臨床分期如下:
(一)前驅期起病緩急不一,大多有低熱或中等熱度,乏力不適,伴有咽痛、咳嗽等上呼吸道症狀,或有納呆,噁心、嘔吐、便秘、腹瀉、腹痛等消化道症狀。神經系統尚無明顯異常。上述症狀持續數小時至3~4天,患者體溫迅速下降而痊癒(稱頓挫型),一部分患者進入癱瘓前期。
(二)癱瘓前期可在發病時即出現本期症狀,或緊接前驅期後出現,或二期之間有短暫間歇(約1~6天),體溫再次上升(稱雙峰熱見於10%~30%患者以小兒為多),出現神經系統症狀如頭痛、頸、背、四肢肌痛,感覺過敏。病兒拒撫抱,動之即哭,坐起時因頸背強直不能前俯,不能屈曲,以上肢向後支撐,呈特殊三角架體態。亦不能以下頦抵膝(吻膝征)。患兒面頰潮紅,多汗,顯示交感神經機能障礙,大多精神興奮,易哭鬧或焦慮不安,偶爾由興奮轉入萎靡、嗜睡。可因頸背肌痛而出現頸部阻力及陽性克氏征、布氏征,肌腱反向及淺反射後期減弱至消失,但無癱瘓。此時腦脊液大多已有改變。一般患者經3~4天熱下降,症狀消失而愈(無癱瘓型)。本期有時長達十餘天。少數患者在本期末出現癱瘓而進入癱瘓期。
(三)癱瘓期一般於起病後3~4天(2~10天)出現肢體癱瘓,癱瘓可突然發生或先有短暫肌力減弱而後發生,腱反射常首先減弱或消失。在5~10天內可相繼出現不同部位的癱瘓,並逐漸加重;輕症則在1~2天后就不再進展。癱瘓早期可伴發熱和肌痛,大多患者體溫下降後癱瘓就不再發展。臨床上分以下幾類。
1.脊髓型麻痹呈弛緩性癱瘓,肌張力低下,腱反射消失,分布不規則,亦不對稱,可累及任何肌肉或肌群,因病變大多在頸、腰部脊髓,故常出現四肢癱瘓,尤以下肢為多。近端大肌群如三角肌、前脛肌等較遠端手足小肌群受累為重,且出現早。軀幹肌群癱瘓時頭不能豎直,頸背乏力,不能坐起和翻身等。癱瘓程度可分為6級:0級(全癱瘓):肌肉刺激時無任何收縮現象;1級(近全癱瘓):肌腱或肌體略見收縮或觸之有收縮感,但不引起動作;2級(重度癱瘓):肢體不能向上抬舉,只能在平面上移動;3級(中度癱瘓):可以自動向上抬舉,但不能承受任何壓力;4級(輕度癱瘓):可以自動向上抬舉,並能承受一定壓力;5級:肌力完全正常。
頸胸部脊髓病變嚴重時可因膈肌和肋間肌(呼吸肌)癱瘓而影響呼吸運動,臨床表現呼吸淺速、聲音低微、咳嗽無力、講話斷斷續續等。體檢可見胸廓擴張受限(肋肌癱瘓)及吸氣時上腹內凹的反常動作(膈肌癱瘓)。若以雙手緊束胸部觀察膈肌動作或手按壓上腹部觀察肋間肌運動,可分辨其活動強弱。膈肌癱瘓時X線透視下可見吸氣時橫膈上抬的反常運動。偶見尿瀦留或失禁(膀胱肌癱瘓)、便秘(腸肌或腹肌癱瘓),常與下肢癱瘓並存,多見於成人。很少發生感覺異常。
2.延髓型麻痹(腦幹型麻痹或球麻痹)
病情多屬嚴重,常與脊髓麻痹同時存在,可有以下表現。
⑴腦神經麻痹:
多見第7、9、10、12對腦神經胺受損。第7對腦神經麻痹常單獨引起面癱,表現為歪嘴、眼瞼下垂或閉合不嚴;軟齶、咽部及聲帶麻痹則因第9、10、12對腦神經病變所致。出現發聲帶鼻音或嘶啞、飲水嗆咳或自鼻反流、吞咽困難、痰液積瀦咽部,隨時有發生窒息的危險。體檢可見軟齶不能上提,懸雍垂歪向健側,咽後壁反射消失,舌外伸偏向患側。動眼障礙和眼瞼下垂見於第3、4、6對腦神經受損;頸無力,肩下垂、頭後傾則見於第11對腦神經受損。
⑵呼吸中樞損害:
以延髓腹面外側網狀組織病變為主。出現呼吸淺弱而不規則,時有雙吸氣和屏氣,呼吸間歇逐漸延長,甚至出現呼吸停頓、脈搏細速和血壓升高(最後下降)。初起表現焦慮不安,繼而神志模糊,進入昏迷,發生嚴重呼吸衰竭。
⑶血管舒縮中樞損害:
以延髓腹面內側網狀組織病變為主。開始面頰潮紅,脈細速不齊,而後轉微弱,血壓下降,皮膚紫紺,四肢濕冷、循環衰竭,患者由極度煩躁不安轉入昏迷。
3.脊髓延髓型較多見,兼有上述兩型的症狀。
4.腦型極少見。可表現為煩躁不安、失眠或嗜睡,可出現驚厥、昏迷及痙攣性癱瘓,嚴重缺氧也可有神志改變。
(四)恢復期及後遺症期急性期過後1~2周癱瘓肢體大多以遠端起逐漸恢復,腱反射也逐漸復常。最初3~6個月恢復較快,以後仍不斷進步,但速度減慢,1~2年後仍不恢復成為後遺症。若不積極治療,則長期癱瘓的肢體可發生肌肉痙攣、萎縮和變形,如足馬蹄內翻或外翻、脊柱畸形等。由於血液供應不良,局部皮膚可有水腫,骨骼發育受阻,嚴重影響活動能力。腸麻痹及膀胱麻痹大多急性期後就恢復,很少留有後遺。呼吸肌麻痹一般在10天內開始恢復,最終完全恢復。極個別需長期依賴人工呼吸器,腦神經受損復元需要一定時日,但很少留有後遺症。
防治原則
簡介
目前尚無特異的治療脊髓灰質炎病毒感染的藥物。對該病的控制主要依賴於疫苗的使用,被動免疫僅用於個別情況。
主動免疫
 服用藥丸
服用藥丸自50年代中期以來,一直採用Salk滅活疫苗及Sabin減毒活疫苗,免疫效果良好,極大地降低了脊髓灰質炎的發病率。Salk疫苗由三型病毒經甲醛滅活後混合製成,肌肉注射,可誘導機體產生中和抗體。其優點是便
於保存及運輸,無減毒株返祖現象,且副作用較少。Sabin疫苗是用減毒變異株製成,採用口服,方法簡便,不但可使機體產生液抗體。還能刺激腸壁漿細胞產生分泌型lgA,對野毒株有消滅作用,從而切斷其在人群中的傳播,因而Sabin 疫苗的免疫效果更好。另外活疫苗病毒排出體外,使接觸者受到感染而獲得免疫。但減毒活疫苗不耐熱,保存及運輸均需冷藏,而且有恢復毒力的危險,在免疫陷人體內易致麻痹。目前世界上大多數國家(包括中國)已將單價脊髓灰質炎活疫苗免疫改為三價活疫苗免疫法,即免疫對象口服三次三價活疫苗糖丸,每次間隔6~8周。其優點是不會漏服,服用次數少,免疫效果好。
被動免疫
用人免疫球蛋白來保護脊髓灰質炎病毒的接觸者。此球蛋白往往含有三型病毒的抗體,及時給予可中和血液中的病毒。被動免疫僅用於做過扁桃腺切除的兒童、未經過免疫接種而又必須接觸脊髓灰質病人的醫務人員和親屬,以及未比免疫接種的孕婦等。免疫效果保持3~5周。
研究進展
經過兩年的辛苦勞作,科學家們首次合成出脊髓灰質炎病毒。這種人造病毒能殺死老鼠,並且很難將它們同自然病毒區分開。目前還不清楚合成類似天花這樣的大病毒來製造生物武器的難度有多大。
脊髓灰質炎病毒是導致小兒麻痹症的罪魁禍首,其基因組是單鏈RNA。感染細胞時,RNA轉譯出一個大分子多聚蛋白,經酶切後形成一組較小的蛋白並向中樞神經系統發動進攻。儘管這個曾經猖獗一時的惡魔已經近乎絕跡了,脊髓灰質炎病毒還在全球實驗室中保留著。
 研究進展
研究進展美國斯托尼·布魯克的紐約州立大學的病毒學家Jeronimo Cello, Aniko Paul和Eckard Wimmer構建了一個近乎完美的病毒複製品。由於RNA化學性質不穩定,研究組先製造了病毒的DNA版本。他們從生物技術公司購買了DNA的短小片段,並在DNA合成公司的協助下將它們連線起來。研究組插入19個標記區分自然病毒與合成病毒,然後利用酶將DNA逆轉為RNA。無論是注入合成病毒還是自然病毒,老鼠都會在感染的一周后癱瘓,研究組在《科學》快訊的電子版發表了這一結論。但是,殺死一隻動物需要1000—10000倍的合成病毒,研究組猜測是一個或多個標記阻礙了病毒的發作。
科學家們對這一研究的技術成果歡呼不已,但也有科學家對其隱含的危機深感憂慮。疾病預防控制中心的James LeDuc說,“化學實驗室的研究人員製造病毒需要保持一絲冷靜”。但是,他並不認為很快就能實現對諸如天花這樣的病毒的重建,這類病毒有超過18.5萬個鹼基對。研究人員說,他們請教過的科學家們督促他們發表這篇文章。正如Cello指出的那樣,“通過把這一結論公布於眾,你們能警示當局……告訴他們生物恐怖主義者都能做些什麼”。
也有人指出,這一發現意味著如果一個替代品能這么簡單地被製造出來,那么消除這個星球上苦難的根源就是不可能的。美國軍隊傳染病醫學研究學院的天花學家Peter Jahrling 說,“它腐蝕了整個根除概念的基礎”。
