
概述
 轉導
轉導轉導(transduction)由噬菌體將一個細胞的基因傳遞給另一細胞的過程。它是細菌之間傳遞遺傳物質的方式之一。其具體含義是指一個細胞的DNA或RNA通過病毒載體的感染轉移到另一個細胞中。
類別
普遍性轉導
 轉導
轉導噬菌體能傳遞供體細菌的任何基因的轉導。鼠傷寒沙門氏菌的P22噬菌體、大腸桿菌P1噬菌體、枯草桿菌轉導的PBS1、PBS2、SP10噬菌體都是普遍性轉導噬菌體。由普遍性轉導產生的轉導子(即接受了噬菌體傳遞的供體細胞基因的受體細胞)不具溶源性,說明轉導噬菌體中不帶有完整的噬菌體染色體,卻帶有噬菌體在繁殖過程中錯誤包裝。
普遍性轉導的供體細菌的基因。根據噬菌體轉導的供體細胞DNA是否整合到受體細胞染色體上,又可將普遍性轉導分為完全轉導和流產轉導。轉導的DNA整合到受體細胞染色體上,並能產生穩定的轉導子的轉導稱為完全轉導;轉導的DNA不整合到受體細胞的染色體上,雖然不能繼續複製,但仍然表達基因功能的轉導稱為流產轉導。在流產轉導的情況中轉導子在每一次細胞分裂時只把噬菌體轉導的DNA傳給兩個子細胞中的一個,所以即使經幾次分裂產生許多細菌,也只有其中的一個細菌細胞得到噬菌體轉導的DNA,這是一種單線遺傳的方式。在同一次轉導中,流產轉導的細胞往往多於完全轉導的細胞。
局限性轉導
噬菌體只能傳遞供體染色體上原噬菌體整合位置附近的基因的轉導。λ噬菌體和φ80噬菌體是大腸桿菌K-12的局限性轉導噬菌體。λ噬菌體只能轉導大腸桿菌K-12染色體半乳糖基因(gal)和生物素基因(bio)等少數基因。φ80噬菌體只能轉導色氨酸基因(trp)、胸腺嘧啶激酶基因(tdk)等少數基因。產生這種現象的原因是由於在溶源化過程中噬菌體總是整合在供體細胞染色體的特定位置上,當溶源性細菌受紫外線等因素誘導後原噬菌體便脫離細菌染色體而進行複製,一部分原噬菌體脫離寄主染色體時帶有鄰近的染色體基因,這些噬菌體便是轉導噬菌體。
(1)低頻轉導(LFT)
誘導溶源性細菌而產生的細胞裂解產物中,除含有正常的噬菌體外,還有極少數(約為10^(-6))部分缺陷噬菌體。用誘導溶源性菌株得來的噬菌體進行轉導時的轉導頻率不過10^(-6) ,稱為低頻轉導。
(2)高頻轉導(HFT)
雙重溶原菌在紫外輻射等因子的誘導下,原噬菌體容易被切割下來,產生等量的缺陷噬菌體和正常噬菌體,該裂解物稱為高頻率轉導裂解物,用這樣的裂解物去感染細菌,將比低頻率轉導裂解物產生多得多的轉導子。
主要途徑
 G蛋白介導的信號轉導途徑
G蛋白介導的信號轉導途徑G蛋白介導的信號轉導途徑
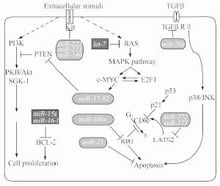
G蛋白可與鳥嘌呤核苷酸可逆性結合。由γ亞基組成的異三聚體在膜受體與效應器之間起中介作用。小G蛋白只具有G蛋白亞基的功能,參與細胞內信號轉導。信息分子與受體結合後,激活不同G蛋白,有以下幾種途經:(1)腺苷酸環化酶途徑 通過激活G蛋白不同亞型,增加或抑制腺苷酸環化酶(AC)活性,調節細胞內cAMP濃度。cAMP可激活蛋白激酶A(PKA),引起多種靶蛋白磷酸化,調節細胞功能。(2) 磷脂酶途徑 激活細胞膜上磷脂酶C(PLC),催化質膜磷脂醯肌醇二磷酸(PIP2)水解,生成三磷酸肌醇(IP3)和甘油二酯(DG)。IP3促進肌漿網或內質網儲存的Ca2+釋放。Ca2+可作為第二信使啟動多種細胞反應。Ca2+與鈣調蛋白結合,激活Ca2+/鈣調蛋白依賴性蛋白激酶或磷酸酯酶,產生多種生物學效應。DG與Ca2+能協調活化蛋白激酶C(PKC)。
受體酪氨酸蛋白激酶(RTPK)信號轉導途徑
受體酪氨酸蛋白激酶超家族的共同特徵是受體本身具有酪氨酸蛋白激酶(TPK)的活性,配體主要為生長因子。RTPK途徑與細胞增殖肥大和腫瘤的發生關係密切。配體與受體胞外區結合後,受體發生二聚化後自身具備(TPK)活性並催化胞內區酪氨酸殘基自身磷酸化。RTPK的下游信號轉導通過多種絲氨酸/蘇氨酸蛋白激酶的級聯激活:(1)激活絲裂原活化蛋白激酶(MAPK),(2)激活蛋白激酶C(PKC),(3)激活磷脂醯肌醇3激酶(PI3K),從而引發相應的生物學效應。
非受體酪氨酸蛋白激酶途徑
此途徑的共同特徵是受體本身不具有TPK活性,配體主要是激素和細胞因子。其調節機制差別很大。如配體與受體結合使受體二聚化後,可通過G蛋白介導激活PLC-β或與胞漿內磷酸化的TPK結合激活PLC-γ,進而引發細胞信號轉導級聯反應。
受體鳥苷酸環化酶信號轉導途徑
一氧化氮(NO)和一氧化碳(CO)可激活鳥苷酸環化酶(GC),增加cGMP生成,cGMP激活蛋白激酶G(PKG),磷酸化靶蛋白發揮生物學作用。
核受體信號轉導途徑
細胞內受體分布於胞漿或核內,本質上都是配體調控的轉錄因子,均在核內啟動信號轉導並影響基因轉錄,統稱核受體。核受體按其結構和功能分為類固醇激素受體家族和甲狀腺素受體家族。類固醇激素受體(雌激素受體除外)位於胞漿,與熱休克蛋白(HSP)結合存在,處於非活化狀態。配體與受體的結合使HSP與受體解離,暴露DNA結合區。激活的受體二聚化並移入核內,與DNA上的激素反應元件(HRE)相結合或其他轉錄因子相互作用,增強或抑制基因的轉錄。甲狀腺素類受體位於核內,不與HSP結合,配體與受體結合後,激活受體並以HRE調節基因轉錄。
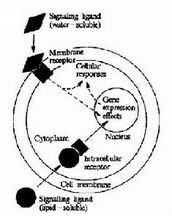
細胞信號轉導
細胞信號轉導是指細胞通過胞膜或胞內受體感受信息分子的刺激,經細胞內信號轉導系統轉換,從而影響細胞生物學功能的過程。水溶性信息分子及前列腺素類(脂溶性)必須首先與胞膜受體結合,啟動細胞內信號轉導的級聯反應,將細胞外的信號跨膜轉導至胞內;脂溶性信息分子可進入胞內,與胞漿或核內受體結合,通過改變靶基因的轉錄活性,誘發細胞特定的應答反應。
媒介
 轉導
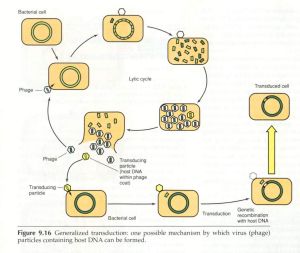
轉導以噬菌體為媒介,把供細菌的基因轉移到受體菌內,導致後者基因改變的過程稱為轉導。當噬菌體在細菌中增殖並裂解細菌時,某些DNA噬菌體(稱為普遍性轉導噬菌體)可在罕見的情況下(約105~107次包裝中發生一次),將細菌的DNA誤作為噬菌體本身的DNA包入頭部蛋白衣殼內。當裂解細菌後,釋放出來的噬菌體通過感染易感細菌則可將供體菌的dna攜帶進入受體菌內。如發生重組則受體菌獲得了噬菌體媒介轉移的供體菌DNA片段。這一過程稱為普遍性轉導。質粒也有可能被包入衣殼進行轉導。不具有轉移裝置的質粒依賴噬菌體媒介進行轉移,轉導可轉移比轉化更大片段的DNA,轉移DNA的效率較轉化為高。
只有溫和噬菌體可進行局限性轉導。當溫和噬菌體進入溶原期時,則以前噬菌體形式整合於細菌染色體的一個部位。當其受激活或自發進入裂解期時,如果該噬菌體DNA在脫離細菌染色體時發生偏離,則僅為與前噬菌體鄰近的細菌染色體DNA有可能被包裝入噬菌體蛋白質衣殼內。因此局限性轉導噬菌體所攜帶的細菌基因只限於插入部位附近的基因。由於局限性轉導噬菌體常缺少噬菌體正常所需的基因,因此常需與野生型噬菌體共同感染細菌後的細菌中複製,這樣才能將攜帶的基因轉移至受體菌,並獲得該段基因所決定的新特性的表達。
套用
套用轉導是細菌的遺傳學研究中的一種常用研究手段。它可以用來在細菌間轉移基因,進行互補測驗,進行基因定位,特別是通過共轉導方法進行基因的精細結構分析。在遺傳工程中可以把所要克隆的基因通過重組DNA技術插入到λ噬菌體的DNA中,然後通過離體包裝方法把它用噬菌體外殼蛋白包裝起來,再去感染寄主細胞以製備基因文庫。
轉導實驗中常用的是λ噬菌體,它能整合在大腸桿菌染色體DNA上的半乳糖基因(gal.17分鐘處)與生物素基因(bio.18分鐘處)之間,因此,它能轉導gal基因又能轉導bio基固。選用溶源性的E.coliK12(λ)gal+為供體菌。由於在此供體菌中λ噬菌體與gal+基因緊密連鎖,因此,當此供體菌受紫外線照射後產生裂解反應,噬菌體被誘發釋放,以一定的比例形成帶有gal基因的轉導噬菌體。當讓這種轉導噬菌體與受體菌E.coliK12Sgal-混合接觸時,帶有供體菌gal+基因的轉導噬菌體能以一定的頻率整合到受體菌的染色體DNA,而使不能利用半乳糖的gal-受體菌轉變成了能利用半乳糖的gal+細菌。
