草原生態
草原生態由地理和氣候條件決定。
草原生態系統主要包括植物、遷徙的野生動物、人類和飼養的牲畜等組成。
草原是地球最大的碳儲庫之一,占地球有機碳總量的33%—34%,是目前人類活動影響最為嚴重的區域,草原生態系統對維持全球及區域性生態平衡有極其重要的作用。
這裡說的是內蒙古大草原,千年來中國人對這片大草原最著名和最準確的描述是白居易的“離離原上草,一歲一枯榮。野火燒不盡,春風吹又生。遠芳浸古道,青翠接荒城。。。”。
當代學者對這片大草原生態系統最準確的描述來自下面的文字:
“首先向各位概述一下中國北方草原生態和社會的一些特點,事實上在中國所謂長城以北是一個乾旱區和半乾旱區的生態系統,實際上是有自身獨特的生態特點的,就表現在自然資源高度的不確定性,主要體現在降水量非常少,可能在有些地區低於年降水量150毫米,而且降水的變率非常大,雨量、降水時間、地點都非常不確定,在這種自然資源高度不確定的情況下,人們如何維持他的生計?在草原區事實上生活著蒙古的遊牧族群,他們在適應這種高度不確定的環境當中有自身的策略,這個策略的核心特點就是流動。這個流動並不是我們很多人所想像的那樣,是一種漫無邊際、沒有目的的流動,事實上在這個看不到邊的蔚藍的草原上有著非常清晰的社會邊界,雖然這種邊界不是固定的,但是它非常清晰,這種邊界是依賴於社會的規範來實踐的,它非常明確的規範著人們的行動。事實上這種流動性不僅體現在遊牧族群能夠在多變的生態條件下靈活應對的這樣一種能力,而且也體現了他們自身的社會組織在不確定的條件下保持秩序和整合的一種能力,可能對現代社會的社會組織方式有很多借鑑的意義。” -荀麗麗(中國社科院)
現在的內蒙古大草原生態已經發生了很大變化,主要是大量農業人口進入請看下圖:
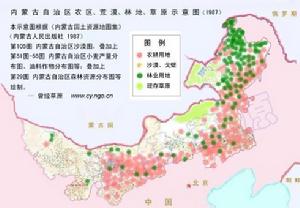 內蒙古天然草原、荒漠、林地、農耕地區位置
內蒙古天然草原、荒漠、林地、農耕地區位置正文
━━━━━━━━━━━━━━━━━━━━━━━━草原概述
草原動植物區系
世界各地草原概述
歐亞大陸草原
北美大陸草原
南美草原
稀樹草原
草原的發生與演替
草原生產力與能流
草原的物質循環
氮循環
磷循環
草原的開發利用
━━━━━━━━━━━━━━━━━━━━━━━━
草原上的生物之間及生物與環境之間的關係。草原是內陸半乾旱到半濕潤氣候條件下所特有的一種自然生態系統類型。一般遼闊無林,旱生多年生禾草占絕對優勢,多年生雜草及半灌木也起顯著作用;通常棲居著各種善於賓士或營穴洞生活的哺乳動物,以及其他動物。
草 原 概 述
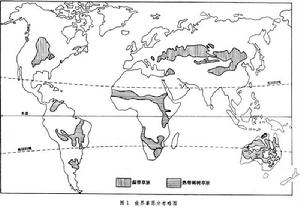
世界草原總面積約2400萬平方公里,為陸地總面積的1/6,大部分用作天然放牧場。草原可分為溫帶草原與熱帶草原兩類。溫帶草原分布在南北兩半球的中緯度地帶,如歐亞大陸草原、北美大陸草原、南美草原等。這裡夏季溫和,冬季寒冷,春季或晚夏有一明顯的乾旱期。由於低溫和雨量較少,草群較低,其地上部分高度多不超過 1米,以耐寒的旱生禾草為主。土壤中以鈣化過程和生草化過程占優勢。分布在熱帶、亞熱帶的草原,其特點是在高大禾草(常達2~3米)的背景上常散生一些不高的喬木,故被稱為稀樹草原。這裡終年溫暖,雨量常達1000毫米以上。在高溫多雨影響下,土壤強烈淋溶,以磚紅壤化過程占優勢,比較貧瘠。一年中存在一個到兩個乾旱
 草原生態
草原生態草原是一種地帶性的生態系統類型。地球上的草原雖然從溫帶分布到熱帶,但在氣候坐標軸上卻占據固定的位置。在寒溫帶,年雨量150~200毫米地區已有大面積草原分布,而在熱帶,稀樹草原卻主要分布在 800~1400毫米雨量的地區,前者的低溫少雨與後者的高溫多雨產生相似的生物學效果。因此,水分與熱量的組合狀況是影響草原分布的決定因素。在地理分布上,草原處於濕潤的森林區與乾旱的荒漠區之間。靠近森林一側,氣候半濕潤,草木繁茂,種類豐富,並常出現島狀森林或灌叢,如北美的高草草原、歐亞大陸的草甸草原以及非洲的高稀樹草原。靠近荒漠一側,雨量減少,氣候變乾,草群低矮稀疏,種類組成簡單,並常混生一些旱生小半灌木或肉質植物,如北美的矮草草原、中國及蒙古的荒漠草原以及蘇聯歐洲部分的荒漠草原等。在上述二者之間為遼闊而典型的禾草草原(圖1)。總的看來,草原因受水分條件的限制,其動植物區系的豐富程度及生物量比森林低,比荒漠高,但與森林和荒漠比較起來,草原動植物種的個體數目以及較小單位面積內種的飽和度是相對豐富的。
草原動植物區系
作為生產者的綠色植物是草原生態系統組成的基礎,草原植物或多或少具旱生特徵。其基本生活型為地面芽植物,尤其是叢生禾草,是草原群落的主要建造者。地下芽植物在草原上也起著重要作用,其中如根莖禾草與根莖苔草,在不少草原地區(尤其是輕質土壤上)起著優勢作用。主要由雙子葉植物組成的各種雜類草,在草原上也非常豐富。如菊科、豆科和薔薇科的一些種,在靠近森林地區即水分條件較好地段繁茂發育,與禾草共同起優勢作用,其根系深,能貯藏水分和養分,故也能分布於乾旱地區。有些雜類草的根系伸至禾草根系層之下才開始分枝,其深度可達禾草根系深度的2倍(圖2)。至於地上芽植物,主要是不高的灌木與半灌木,在草原上多少起顯著作用,尤其在接近荒漠地區,它們的 草原生態
草原生態生態學家常按高度把草原上占優勢的多年生草本植物分為 1米以上的高草、30~90厘米的中草和30厘米以下的矮草。高草主要分布在水分條件較好的熱帶、亞熱帶稀樹草原以及溫帶半濕潤地區,多為中旱生植物甚至中生植物;矮草分布在靠近荒漠一側的乾旱地區,形成矮草草原和荒漠草原,多為典型的旱生植物和強旱生植物;中草的分布介於二者之間,為典型草原的主要成分。
根據草原植物對溫度的反應,可分出耐寒型和喜暖型兩大類。耐寒型多分布在中溫帶、寒溫帶及高寒山地草原群落中,以早熟禾系的禾草為主,如針茅、羊茅、早熟禾、冰草、隱子草等屬的一些種,它們多為呌植物;喜暖型則分布在熱帶、亞熱帶稀樹草原以及溫帶草原的暖溫帶地區,以黍系禾草為主,如黍、狼尾草、芒須草、扭黃茅、萑稗、高粱等屬的一些種,它們多為C4植物。
不同地區草原植物種類的豐富程度不同。生態條件越適宜,種類就越豐富,群落結構也越複雜,有地上及地下亞層的分化。反之,生態條件越嚴酷,種類就越簡單,群落結構也越簡化。如中國大興安嶺兩側的草甸草原群落,每平方米約有種子植物20~30種或更多;廣大的典型草原,每平方米約15~20餘種;而乾旱的荒漠草原,每平方米僅12種左右。在種的飽和度低、群落結構趨於簡化的情況下,地上部分常常不能鬱閉,覆蓋度多在30%以下,但其地下部分卻是鬱閉的。草原植物地上生物量與地下生物量的比值隨生境的變劣而降低。
在草原群落中起建群作用的植物只是很少數,絕大部分植物為伴生種。如中國草原地區共有種子植物4000種左右,但建群植物只有45~50種,其中主要是禾草,約占全部建群種的75%。由於建群植物的形成歷史與生態性質不同,其分布有明顯的地域分異,從而形成不同的草原類型。
草原動物區系也很複雜,最引人注目的是大型植食動物,如非洲的斑馬、瞪羚、長頸鹿,美洲的野牛,大洋洲的有袋類、麋鹿,歐亞大陸草原的野驢、黃羊等以及牛羊等家畜。此外還有數量眾多的其他類群,它們大部分為植食性。植物性食料雖易獲得,但營養價值不及動物性食料,必須攝取更大的量才能滿足營養要求,因而植食動物在形態上和行為上有一系列的適應。駱駝、袋鼠、長頸鹿、鹿及牛、羊等各自獨立地發展了反芻習性。這一特性可以使它們的採食時間縮短,到達安全地區後再將所進食物反芻咀嚼,從而減少了被捕食的機會。營地上生活的草原動物常以快速賓士保證其安全,常可見到一些善跳善跑類群。善跳的如袋鼠、跳鼠、兔與跳兔,善跑的如羚羊、黃羊、鹿等。它們體輕腿長,異常敏捷,在空曠的草原上能將捕食者遠遠地拋在後面。鳥類中的鴕鳥、鴯鶓等也屬於善跑的一類。有些小型哺乳動物(如嚙齒類)則營地下穴洞生活,以適應草原氣候的季節變化和逃避捕食者。它們挖掘洞穴,使土壤混合與疏鬆,在草原群落的形成與演替中起著很大作用。至於大型哺乳動物如象與野牛等,則以其強大的體型為自衛手段。草原動物的視力多敏銳,一些小型哺乳動物在觀察周圍情況時,常會遇到草叢的障礙,於是其中的一些種類用後腿坐起來窺伺周圍環境。多種多樣的草原肉食動物,從大型的獅、豹、獵豹到狼、狐、獾及鷹等,它們可以抑制某些草食動物種群數量的大發生,從而維持草原生態系統的穩定與平衡。無脊椎土壤動物的數量更多,它們在草原生態系統中也起著顯著的作用。
世界各地草原概述
歐亞大陸草原 地球上最遼闊的溫帶草原區域,自歐洲多瑙河下游起,呈帶狀往東延伸,經匈牙利、羅馬尼亞、蘇聯、蒙古人民共和國,直至中國東北平原,然後轉向西南,經內蒙古高原、黃土高原達青藏高原的南緣,綿延 8000餘公里(圖3)。這一區域環繞在亞洲荒漠區的外圍,東西兩側與溫帶落葉闊葉林相接,北連寒溫帶針葉林。
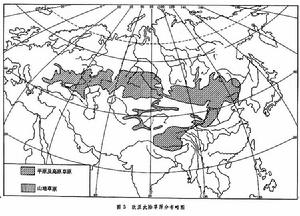 草原生態
草原生態在歐亞大陸草原範圍內,水熱條件在一年中的分配狀況各地有較大差異。西部為地中海型氣候,冬春溫和多雨,夏季炎熱乾旱。草原生物於春季繁茂生長,夏季往往進入休眠狀態,草群中發育了大量春季短命植物和類短命植物。草原區東部為太平洋季風氣候,冬季寒冷少雪,夏季炎熱多雨,水熱同季,植物生長呈單峰型,夏季最為繁茂,缺少春季短命植物及類短命植物。青藏高原降水分配也為季風型,但氣溫終年較低,尤其是生長季氣溫低,因此這裡發育了適應於高寒氣候的動植物類型,草原中出現墊狀植物層片。
除了上述區域差別之外,因水熱組合狀況不同而引起的地帶分異也很明顯。在歐亞大陸草原區內,從靠近森林一側向荒漠區推近時,年降水量逐漸降低,熱量漸趨增加,從而引起草原的一系列變化。主要是有效水分越來越少,土壤淋溶程度逐漸減弱,鈣積層漸趨於地表,植物高度、密度與種的數目漸趨降低等,動植物區系組成也發生相應的更替,出現森林草原、典型草原與荒漠草原3個自然帶的分化。
森林草原帶 動植物區系豐富,草群密茂,並有島狀樺、楊林及櫟林出現。草群下發育了肥厚的黑土、黑鈣土及黑壚土。其建群植物西部為約翰針茅、狹葉針茅,蒙古高原東部和中國東北部為貝加爾針茅、中國濱麥及線葉菊,黃土高原為牡羊草,草群中混生相當豐富的中生性雜類草。大型草食動物主要有駝鹿、鹿類、麅、原羚等。由於這一地帶氣候適宜,土壤肥沃,已大部開墾為農田,為春小麥的主要產區。
典型草原帶 比森林草原帶更偏向內陸,具典型的半乾旱大陸性氣候,發育了栗鈣土。草原植物組成簡單,以旱生叢生禾草尤其是針茅屬占絕對優勢,當針茅開花季節(6~8月),其銀色的長芒隨風波浪起伏,一望無際,十分壯觀。西部以幾種羽狀芒針茅占優勢,如列興針茅、針茅、紫針茅等;東部建群植物以光芒組的幾種針茅為主,如大針茅、克氏針茅、長芒草以及糙隱子草、冷蒿等,草群中缺少中生性雜類草。典型草原上常見的草食動物有黃羊、高鼻羚羊以及黃鼠、鼠兔、旱獺等。目前,典型草原地帶除局部開墾為農田外,大半保持自然狀態,為放牧畜牧業的主要基地。
荒漠草原帶 從典型草原到荒漠的過渡地帶。年降水量僅120~180毫米,而蒸發量卻數倍於此值,是草原區內最乾旱的地區。草群低矮稀疏,草高僅20厘米左右,覆蓋度很低,這為一年生植物的發育提供了空間。每逢多雨年份的雨季,生長起繁盛的一年生植物層片,為草原增添了不少色彩。荒漠草原的群落組成極為特化,如須芒組的小針茅、亞菊等屬的小半灌木以及不少嚙齒類動物均為這一地帶所特有。主要建群植物有新疆針芒、戈壁針茅、短花針茅、蓍狀亞菊、女蒿、鹼韭以及青藏高原的青藏苔草等。荒漠草原的主要草食動物為沙鼠、短耳倉鼠、高鼻羚羊等。由於蒸發強烈,土壤中的鈣及易溶性鹽類積累於地表,腐殖質含量低,為棕鈣土。這裡無灌溉即無農業,但可用作良好的放牧場。
北美大陸草原 分布也極為遼闊,北從加拿大南部起,經美國中部達墨西哥灣;西起落基山東麓,東至美國五大湖區西岸,南北延伸約3700公里,東西寬約700~800公里(圖4)。北鄰針葉林帶,南部逐漸過渡到稀樹草原,東接落葉闊葉林,西為落基山山地針葉林及蒿類草地。
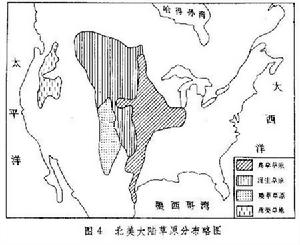 草原生態
草原生態北美草原的分布格局與歐洲大陸有所不同。這裡雨量從東往西減少,東部邊緣年降水量達 750(北段)~1000毫米(南段),中部500毫米上下,西部降至400毫米以下。與此相應,優勢植物的高度從東往西逐漸降低,種類組成也發生明顯變化,出現3個地帶的依次交替:高草草原、混生草原與矮草草原。從北而南氣溫逐漸增高,最冷月(1月)平均氣溫北部低於-20℃,而南部高於0℃,反映在草原組成上也發生明顯變化,開花遲的喜暖種在南方占優勢,而在北方則生長耐寒種類。
高草草原 發育在肥沃的黑土上,原始狀態下以須芒草屬植物占優勢。已多被開墾,種植禾穀類作物及多年生牧草。
混生草原 特點是高草與矮草混生,顯然是一過渡帶。
矮草草原 從混生草原帶往西南,高草逐漸消失,過渡到矮草草原。這裡以野牛草及格蘭馬草等矮禾草占絕對優勢,草群高度30厘米左右,根深達 1米。種的豐富度明顯降低,1平方公里僅20種上下,雜類草的數量下降,但混生一些矮灌木、半灌木及肉質植物,如蒿及黃茋等屬的一些種以及肉質化的多刺仙人掌。
動物區系 北美草原曾經棲息過豐富的動物區系,如古象、乳齒象、古駱駝、野牛、駝鹿等植食動物和以它們為食的美洲豹、美洲獅等貓科肉食動物,但大多已經滅絕,只有野牛與駝鹿由於印第安人家養而存活下來。目前,牛、馬、羊等家畜取代了原來的野生動物。
南美草原 主要分布在阿根廷中東部平原,約處於南緯32°~38°之間,面積約50萬平方公里。草原區北部降水量達1000~1250毫米,是所有溫帶草原中降水最高的,往西南雨量漸少,至草原的西南邊緣,降水量為500毫米左右。這裡氣溫較高,蒸發強烈,氣候相當乾燥。當降水量進一步降低,或溫度進一步升高時,草原便為旱生灌叢或稀樹草原所代替。北部輕質土壤上出現具刺朴樹組成的島狀林,優勢草原群落主要由奈西針茅和牡羊草等禾草組成,混生相當多的雜類草;目前原始群落殘存無幾,大部已被農田代替。西部及西南部生長了以南美針茅和三歧針茅為主的大型叢生禾草草原,幾乎很少雜類草。南美草原上的畜牧業僅有400年的歷史,但近一個世紀以來的過度開墾和重牧使生產力持續降低,植被組成也多為引入成分所代替。
稀樹草原 主要分布於非洲、大洋洲及南美洲的熱帶、亞熱帶半乾旱地區。在大洋洲叫scrub,巴西叫cam-pe,南美北部叫llano。後來,南美北部的llano被西班牙人叫做Savanna(稀樹草原),此後就把類似的草地類型都稱之為稀樹草原。
非洲的稀樹草原 分布於赤道雨林的南北兩側,介於北緯15°和南緯25°之間,形成東西延伸的帶,北側頻寬400~550公里,東西長約5000公里;南側頻寬200公里,東西長約2500公里。總面積80多萬平方公里。優勢植物為須芒草族及黍族的高大禾草。散生樹木以具刺的金合歡屬為主,也有合歡亞科的其他屬以及另外一些科的耐旱樹木生長,多具刺。在靠近熱帶雨林一側分布了以狼尾草屬和白茅屬為主的高稀樹草原,往北或往南,隨乾旱程度的增加,草群變低矮,以菅草最為豐富。
稀樹草原植物的生長期決定於乾旱期的長度,一般為120~190天。土壤高度淋溶,以磚紅土壤化過程占優勢。分解過程快,腐殖質積累量低。經常的火災對保持稀樹草原的穩定起著一定作用;但同時也消滅了對火敏感的種類,促進耐火種類的發展,從而成為稀樹草原演替的動力。
非洲稀樹草原的動物區系豐富,其中很多為非洲所特有,如非洲象、長頸鹿、斑馬、非洲野牛、斑鬣狗、非洲野狗、獴等。此外還有很多小型哺乳動物如嚙齒類,數量眾多的鳥類(如著名的鴕鳥、鷹等);以及大量的昆蟲和其他無脊椎動物。這些動物具有相當高的生物量,如對東非 1.5平方公里稀樹草原的估計,有蹄類的生物量每公頃達50千克,小哺乳動物4~5千克,肉食動物8.5千克,無脊椎動物的生物量每公頃可達100千克。
南美稀樹草原 以委內瑞拉的 llano面積最大,位於安第斯山脈奧里諾科河之間,優勢植物為黍屬禾草,而具刺樹木為高大的仙人掌所取代。這裡多被開墾,已很難找到原生植被。
大洋洲稀樹草原 也有相當大面積,這裡仍存在金合歡但卻沒有具刺的樹和灌叢,散生樹木的優勢樹種是桉樹,優勢草本植物為高大禾草Astrebla屬等,雨季繁茂生長,旱季乾枯,營養價值很低。特有動物有袋鼠、樹熊、袋狼等,其中袋狼可能已近於滅絕。
草原的發生與演變
草原的發生至少可追溯到距今 700萬年之前的中新世。從中新世起到第四紀中期,中國地殼曾發生一系列巨大變動。喜馬拉雅山、崑崙山、天山、阿爾泰山和青藏高原先後在西部隆起,同時東部臨海部分升起許多山地,蒙古高原和黃土高原也在這時形成。地形的升起阻擋了北大西洋和印度洋濕氣流東上,同時也減弱了太平洋濕氣團的西進,使中國北部日趨寒冷、乾旱,促成乾旱、半乾旱區的形成。中新世時期,中國乾旱區大致為稀樹草原景觀,僅在山地開始形成草原群落片段。到中新世末,形成於山地的草原群落下降到平地,並往東侵移。第四紀時期的冰期與間冰期的交替,促進了南北之間以及山上與山下之間植物的交流,於是逐漸奠定了現代草原的輪廓。青藏高原的草原群落形成最晚,大概在晚更新世(距今2~10萬年),才形成目前的景觀。隨著草原植被的形成也出現了以草原植物為食,生活在草原環境的特定動物類群。
草原生產力與能流
一般說來,由於受到水分條件的限制,草原的初級生產力在所有陸地生態系統中是中等或中等偏下的。據聯合國的資料,北美草原地上生物量為每平方米 263~1062克,年淨生產量為每平方米 506~1425克。中國草原地上部分生物量,草甸草原約每平方米為200~500克乾物質,典型草原每平方米為 75~200克,荒漠草原僅每平方米為 20~100克。中國草原植物年生產量的估計值為每平方米50~1250克,其上限與北美、蘇聯草原相似,而下限較低,這是因為中國的荒漠草原氣候條件比較嚴酷所致。蒙古荒漠草原生產量的下限為每平方米49.8克,與中國相似。
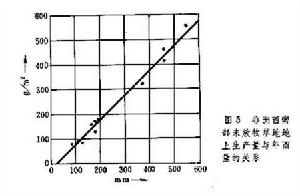 草原生態
草原生態草原初級生產力與水分條件有密切關係。1964年在非洲的研究發現,稀樹草原地上部分淨生產量與年降雨量呈線性相關(圖5)。溫帶草原也存在類似規律。還進一步發現,植物的淨光合作用與葉子及土壤的水勢呈線性關係。因此,從森林草原至荒漠草原,隨雨量的減少,初級生產力有規律地降低。此外,在供水充足的條件下,在年平均氣溫-2~26℃之間,溫度越高則生產力亦越高。因此,稀樹草原的生產力顯著比溫帶草原高,平均達1600克/(米2·年),最高可達4000克/(米2·年)。
在生長季草原植物的光能利用率大約為0.1%~1.4%,沿水分梯度而增減。1967年北美高草草原的估算為1.2%,而1979年估算為0.67%~1.09%。聯合國在北美洲測定這一數值為0.12%~1.43%。蘇聯的草甸草原達1.32%,而蒙古荒漠草原尚不及0.1%。
草原初級生產所創造的有機物質為各種植食動物所取食。草原的植食動物多種多樣,它們以不同方式採食不同植物以及同種植物的不同部分。植食動物又為肉食動物所食。如鼠被鼬吞食,而鼬被鷹食等等。肉食動物的獵取對象也常常是專化的,捕食者與被食者之間互相制約,組成了複雜的食物網。
草原生態系統中,植物所固定的太陽能量沿食物鏈傳遞,逐級減少,所構成的生態金字塔可以美國科羅拉多的矮草草原為代表(圖6)。聯合國在科羅拉多矮草草原上測定,該地生長季內有效太陽輻射總量為每平方米5×105千卡,綠色
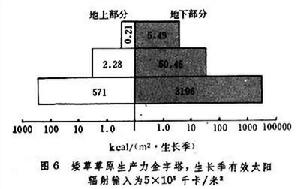 草原生態
草原生態在草原生態系統中,大部分能量沿捕食食物鏈通過,但碎屑食物鏈也很重要。在原始情況下,草原上凋落物的量相當可觀。通常地上凋落物約為地上部分現存量的20%~50%,而地下部分凋落物可能還大於地上部分。在溫帶草原上,地上枯死部分的分解速率約為兩年,地下部分則需 4年左右。凋落物的積累速率大致與分解速率相當,所以在天然草原上常保留著明顯的凋落物層。熱帶稀樹草原的分解速率比溫帶草原快得多,因此,這裡一般看不到凋落物層,土壤中也很少腐殖質的積累。
草原生態系統中分解者的生物量相當高,往往超過消費者的生物量。據加拿大南部馬特多地區1968年測定數字,該地草原植物量為每平方米434克,30厘米土層內土壤微生物量達每平方米254克;1970年另一次測定,相應數字為每平方米622克與328克。當然在不同地區,這一數值很不相同,北美矮草草原的數值就低得多,如據波尼地區測定,30厘米土層內真菌生物量為每平方米38克,細菌為27.6克,總計為每平方米65.6克(1979)。植物凋落物中約有1/3能量轉移給微生物。圖7
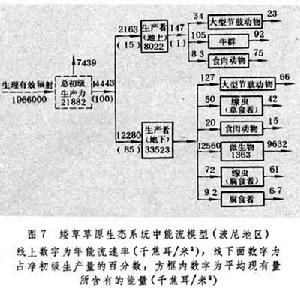 為波尼地區的能流模型。該地區植物生長期內 (154天)的太陽輻射總量為每平方米4158000千焦耳,生理輻射量為每平方米1966000千焦耳,植物淨生產量約固定總輻射量的0.34%,生理輻射量的0.73%。植物所固定的能量約15%分配於地上部分,而85%分配於地下部分(在水分條件較好的馬特多,相應數字為43%與57%)。植物地上部分能量約有 6.7%流入消費者體內,其餘大部分隨枯死部分進入凋落物中。植物地下部分的能量大部分轉入土壤生物,主要是進入土壤微生物。由於土壤微生物的能量不僅僅得自植物地下部分,因此它的流通總量高於當年流入植物地下部分的總量。
為波尼地區的能流模型。該地區植物生長期內 (154天)的太陽輻射總量為每平方米4158000千焦耳,生理輻射量為每平方米1966000千焦耳,植物淨生產量約固定總輻射量的0.34%,生理輻射量的0.73%。植物所固定的能量約15%分配於地上部分,而85%分配於地下部分(在水分條件較好的馬特多,相應數字為43%與57%)。植物地上部分能量約有 6.7%流入消費者體內,其餘大部分隨枯死部分進入凋落物中。植物地下部分的能量大部分轉入土壤生物,主要是進入土壤微生物。由於土壤微生物的能量不僅僅得自植物地下部分,因此它的流通總量高於當年流入植物地下部分的總量。 草原的物質循環
物質循環是草原生態系統的另一重要機能。
氮循環 固氮微生物對草原生態系統氮的輸入起重要作用,其中共生固氮菌,特別是豆科植物的根瘤菌更為重要。在適宜的生長條件下,根瘤菌在人工草地上的固氮量達每年每公頃100~300千克(1970),高產苜蓿地上甚至可達每年每公頃 670千克(1965)。但多數天然草地上豆科植物較少,如中國及北美草原上豆科植物比例一般不到草群總重量的10%,有時甚至不到1%,因此,這裡共生固氮菌的固氮量遠低於上述數字。即使如此,它們的固氮量仍是可觀的。除豆科植物外,草原上還有一些植物能與根瘤菌共生,如盧多維奇氏蒿、纖細仙人掌等。非共生固氮菌在草原氮素輸入中也起一定作用,一般固氮量為1~2千克/(公頃·年);藍藻的作用不大;而通過降水輸入的氮約為3~4千克/(公頃·年)(1980)。大致講來,植物地上部分氮素周轉率至少在50%以上,而地下部分均為19%。研究表明,植物體的氮素只有一部分流入腐殖質,另一部分流入微生物,還有相當一部分以無機氮形式釋放出來(ƒ2,ƒ4)。1980年估計,北美矮草草原上從植物體進入土壤腐殖質的氮不超過10%,而腐殖氮的礦化率每年僅為 0.2%。如波尼附近草原上的植物每年需氮量約每年每平方米7克,其中僅1.8克來自腐殖質,5.2克直接來自凋落物。
氮素可以通過動物移動,植物產品的收割、揮發、淋溶等方式流出生態系統而損失。一般情況下,草原上氮的損失量要比農田小得多,因為在乾旱、半乾旱地區,通過淋溶而損失的氮素微不足道。但在過牧地或施氮肥後,氨態氧可能通過揮發而大量損失。1970年有報導說,排泄在草原乾燥土壤上的牛尿中,90%的氨態氮被揮發掉。野生動物的移動對草原養分循環的影響不大,一般家畜攝取養分的80%~90%以糞便等形式返回土壤,它們帶走的營養物質通常不超過其攝取量的10%(1956)。總之,在自然狀態下氮素的流出量不大。常常是流入量略大於流出量,即土壤庫中的氮素逐年積累。但另一方面,系統內氮素的年輸入量遠不能維持植物年生長對氮素的需求量,如北美矮草草原上植物體所含氮素量達每年每公頃 428千克(1977),這可能是當地氮素年輸入量的10倍以上。可見,已存在於系統中的氮素的再循環對草原是重要的。據同一地點測定,0~36厘米土層中的氮素達每公頃3725千克,幾為植物體氮素的9倍,這說明參與循環的氮素僅為系統內總氮量的一小部分。
值得指出的是,人為地從草原上取走大量動植物產品,便可使草原的氮素循環失去平衡。例如中國草原區東部的天然割草場,常常由於連年刈割而導致產量、質量持續降低。在這種情況下,則須通過施氮肥或種植豆科牧草以增加草原的氮素輸入,保持氮素平衡。開墾原始草原也會使土壤中有機物質的分解加速,收割和水土流失又帶走大量的氮。因此,開墾草原實際上是在開採土壤中的氮貯庫,因此必須通過增施氮肥來維持氮素平衡。
磷循環 草原土壤中磷的總量以及有機磷對無機磷的比例,與氣候條件尤其是氣溫有密切聯繫(1961)。如在美國草原,從南往北隨著氣溫的降低,土壤表層15厘米內總磷量呈現有規律的增高。此外,在總磷中可利用的有機磷數量常常超過無機磷,這點與耕作土壤迥異。看來,在草原的磷循環中,有機磷的轉移起著重要作用。
植物吸收的磷僅為土壤中可利用磷的一小部分。植食動物所攝取的磷,約有60%以上通過糞便排出,歸還土壤,通過動物所移走的磷不到動物吸收總量的10%(1967)。大部分磷在有機物殘體中長期保存下來。土壤中有機磷的礦化與有機氮和碳的礦化有密切關係。1977年估計,在北美草原上,磷的礦化速率約為每平方米3~6.4克。分解者對磷的攝取幾乎5倍於植物吸收量,有機磷的礦化量與磷的總攝取量接近平衡,因此當生長季結束時土壤中有效無機磷的含量幾乎與生長季開始時相等。
草原的開發利用
以單位面積動物量計算,草原在所有陸地生態系統中名列前茅。早在舊石器時代,人們就大量捕殺非洲稀樹草原和歐亞大陸草原的野生動物;當人類第一次越過白令海峽移入北美時,那裡草原動物也遭到同樣的命運。在最近的一次冰期,僅非洲和北美就有61個大型草原哺乳動物的屬被消滅,當然這主要由氣候的重大變化引起,但人類的捕殺也無疑起了一定作用。當人類學會使用武器之後,摧毀草原野生動物的速度加快了。1870年前後,北美草原上還有千萬頭野牛到處遊蕩,隨著對美國西部的開發,30餘年間已把野牛消滅殆盡。現在,這種動物只能在國家公園內才能見到。歐亞大陸草原上的野馬已近絕跡,野驢、野駱駝也已很少。
人類大量捕殺草原野生動物的同時,又把一些對自己有用的動物馴養起來。逐漸地,家養動物代替大型野生動物成為草原上主要的消費者。在原始狀態下,草原生態系統中的食物鏈比較長,食物網也比較複雜。各種動物之間以及動物與植物之間通過相互制約和自然調節而保持穩定與平衡。一旦家畜代替了野生草食動物,種類貧乏了,食物鏈縮短了,草原生態系統變得比較脆弱。當然,放牧畜牧業較之狩獵是人類社會的一大進步,也給人類提供了更多的動物性產品。但在人口壓力日漸增加的今天,不少地區無限制地利用草原,不考慮草原的生產力而盲目增載入畜量,結果使草原生態系統入不敷出,破壞了生態平衡,導致草原生產力的持續降低。因此,在放牧利用條件下,應以草定畜,把牲畜數量保持在能使草原正常更新的範圍內,據試驗,當牲畜採食量長期超過地上產量的50%時,就會引起產量降低,草質變壞,導致經濟收入的降低,只有在這一限度內實行輕牧,報酬才是最高的。因此應在研究不同草原類型的生產力及其動態的基礎上制定適宜的載畜量,以保證草原的更新、保持草原的生態平衡。同時,可通過培育牲畜良種以及加快周轉等措施,增加單位草原面積的畜產品。
除放牧利用外,今天世界草原的很大一部分已被農田所代替。如北美的高草草原目前已開墾殆盡,成為美國玉米和小麥的主要產區;在匈牙利,昔日的“無林草地”目前幾乎皆成為農田;南美東部的草原亦殘存無幾,大部為農田和人工草地所取代;蘇聯草原也進行了大面積墾植。在中國,黃土高原早在千年前已被徹底開墾;鄂爾多斯草原也在幾百年前開墾殆盡,這些地區開墾之後,由於作物單一和粗放經營,水土流失嚴重,土壤肥力迅速消耗,不少地段已成為不毛之地,是中國土地單產最低的區域之一。天然草原的土壤質量較高,尤其當禾草中混生適當比例的蟲媒傳粉的雜類草時,可形成肥力極好的土壤類型,草原黑土與黑鈣土便是地球上最肥沃的土壤。草原轉為農田,消除了天然禾草與多種雜類草,代之以一年生作物,後者腐殖質形成作用很低,而且其產品被大部分取走。因此,草原開墾後若不通過施肥來補充養分,則土壤肥力很快降低。如中國東北平原的黑土,原始狀態下腐殖質層厚達50~70厘米,有機質含量達6%~15%,開荒後4~5年內產量很高,若不施肥,15年後僅及前5年產量的一半,40年後肥力基本耗竭。至於條件較差的草原,這一過程更短得多。可見,草原的開發必須伴隨恢復肥力、保持生態平衡的一些措施,否則將導致資源枯竭。目前草原開墾後保持養分平衡的辦法有幾種:一是輪歇,如美國西部,有些草原開墾後種小麥,呈條帶狀間隔種植,種一年歇一年,條帶間互相輪換。這不但對恢復地力有益,而且對保持土壤水分也起一定作用。二是輪作,作物與苜蓿多年生牧草輪流種植,藉助豆科牧草固定氮素,從而保持土地資源的生產力。三是對已墾農田進行灌水、施肥等耕作措施,不斷施入補加能量。進行集約經營,保持高的生產力。一般地說,施入的補加能量越多,生產力也越高。但近來不少地區發現,當生產力達到一定水平之後,即使繼續投入高額補加能量,生產量的增加也越來越少,而且土壤性狀亦趨變壞。因此,利用輪作維持土壤養分平衡的辦法日益受到重視。中國草原區的農田,過去開墾後廣種薄收,連續單一種植,只取不還,只用不養,結果不少地區土壤肥力及生產力持續下降。今後,草原的利用、改造須考慮到資源的持續利用。在開墾時,就要設計出保持生態平衡的有效措施。
參考書目
中國植被編輯委員會:《中國植被》,科學出版社,北京,1980。
A.I.Breymeyer,G.M.van Dyne, Grassland,S ystems Analysis and Man,Cambridge University Press,1980.
R.T.Coupland, Grassland Ecosystems of the World,Cambridge University Press,1979.
Helmut Lieth, Primary Production of the Major Vegetation Units of the World.Primary Productivity of the Biosphere, Springer-Verlag New York Inc.,1975.
Makoto Numata, Ecology of Grasslands and Bamboolands in the World, VEB Gustav Fischer Verlag,Jena,1979.
