植物水分生理 植物生理學 的一個重要分支,研究和闡明水對植物生活的意義、植物對水的吸收、水在植物體內的運輸和向大氣的散失(蒸騰作用 ),以及植物對水分脅迫的回響與適應。地球表面上降水量 和蒸騰需求分布極不均勻,不同地區乾燥度(蒸騰需求與降水量之比)相差懸殊,植物種的水分生理特點和抗水分脅迫能力,對決定其地理分布和生產力 起重要作用。在農業 上主要關心的是植物的產量和品質。灌溉水充裕時,追求以合理灌水來使產量最高;降水不足、灌溉有限時,則單位水量所能給出的產量最高、即水的經濟利用效率最高更為重要。這在旱作農業中制定各項農業措施(包括品種的選擇)中,是主要的目標。水和植物的生活 植物生理學 原生質 含水量為80~90%,其中葉綠體 和線粒體 含50%左右;液泡 中則含90%以上。組織或器官的含水量隨木質化程度增加而減少,如瓜果的肉質部分含水量可超過90%,幼嫩的葉子為80~90%,根為70~95%,樹幹則平均為50%,休眠芽約40%。含水最少的是成熟的種子 ,一般僅10~14%,或更少。代謝旺盛的器官或組織含水量都很高。原生質只有在含水量足夠高時,才能進行各種生理活動。各種生化反應都須以水為介質或溶劑來進行。水是光合作用 的基本原料之一,它參加各種水解反應和呼吸作用 中的多種反應。植物的生長,通常靠吸水使細胞伸長或膨大。膨壓降低,生長就減緩或停止。如:曇花一現,就是靠花瓣快速吸水膨大、張開;牽牛花 清晨開放,日光曝曬後失水捲縮。某些植物分化出特殊的器官,水分進出造成膨壓可逆地升降,使器官快速地運動。如水稻葉子在空氣乾旱、供水不足時,泡狀細胞失水,使葉片捲成圓筒狀,供水恢復後重新展開;氣孔的運動是通過保衛細胞因水分情況變化而脹縮來實現的,從而調節水分散失速率,維持植株水分平衡。反之,有些器官只有失水時才能完成某些功能。如藤蘿的果莢,只有乾燥時才能爆裂,使其中的種子迸出;蒲公英 的種子成熟、失水後才會脫離母體,隨風飄蕩。水的特有的理化性質給植物帶來一些好處。水的汽化熱(20℃時為2 454J/g)與比熱〔4.187J(g℃)〕特別高,有利於發散植株所吸收的輻射熱;避免體溫 大幅度上升。水的表面張力、內聚力 及與一些物質間的吸附力 在植物體內運輸中有重要意義。水能透過可見光和紫外光 ,使日光能透射到葉綠體上供光合作用 之用,或被光敏素 等吸收,引起光形態發生效應。水分子的極性造成了多種化合物的水合狀態 ,並使原生質親水膠體得以穩定。
陸生植物按其體內水分隨環境乾濕而變化的程度,可分為變水型和恆水型兩大類。變水型植物如地衣、旱生苔蘚和幾十種維管植物 (其中包括少數被子植物)。它們的細胞小而無液泡。環境乾燥時,它們的含水量也隨之迅速減少。大部分變水型植物在乾燥時細胞均勻地皺縮,原生質凝膠化,進入隱生狀態,但原生質結構不受破壞;一旦獲得水分,還能迅速恢復代謝和生長。恆水型植物成熟細胞中有貯存水分的大液泡,在環境水分短暫變化時,可對細胞含水量變化起緩衝作用;有角質層 阻止水分蒸發,氣孔也可隨時關閉;有龐大的根系和特化的輸導組織以便從土壤中吸收水分輸送給地上部。這些特點使恆水型植物能維持體內水分平衡,控制含水量在一定限度之內。幾乎全部高等植物都是恆水型的。植物水分生理的主要內容是恆水型植物的水分代謝。
植物細胞的水分關係 水分子 自由水 是可以移動的。生理上活躍的組織中,大部分水(包括液泡水)是自由水。束縛水 是通過氫鍵吸附於細胞中特別是膜上的蛋白質 、多糖之上的水分子,成半晶體 排列,密度比液態水大。細胞壁微纖絲的纖維素 和多糖膠體表面上,也有束縛水膜存在。細胞中的水分分別存在於以下幾個區隔之中:①細胞壁以及細胞壁間物質,它們由親水性的纖維素和果膠質組成。整個植株的細胞壁 除根部被凱氏帶隔開以外都彼此連線,形成質外體,水分可以在其中自由流動。②細胞質 ,其外由細胞膜(質膜)與質外體隔開,其內以液泡膜與液泡隔開。③細胞質中的各種細胞器,如核、線粒體、葉綠體等,各自有膜包圍。④液泡。各個區隔之間的膜都允許水分子 通過,水透過膜的脂肪層時遇到的阻力較大。水通過的方向取決於膜兩側水勢的高低。由於各種膜都是差透膜,水中溶解的離子以及糖、有機酸等有機化合物 分子的透過性各不相同,因而當一個區隔中幾種物質相互轉化(例如大分子的澱粉或蛋白質轉變為小分子的單糖、雙糖、有機酸或胺基酸)時,其滲透勢會發生很大變化,從而引起水的移動。植物細胞滲透勢的變化可以調節水分關係。例如氣孔通過調節保衛細胞鉀的進出來改變滲透勢,從而提高或降低膨壓,使氣孔開放或關閉。
整體植物的水分代謝 植物水分生理 大氣 形成一個連續的系統,稱為土壤-植物-大氣連續系。與電學 中由電阻組成的線路作類比,可以把水自土壤到大氣所經歷的路徑的各階段看作一系列串聯的阻力,通過這個途徑的水的通量(F)是推動力即水勢差(ΔΨ)被阻力(r)除所得的商。即某一段的水勢差 (或稱水勢降)越大,則它的阻力也越大。從折線可以粗略地看出各段間的水勢差,從而可以估計各段的阻力。各段阻力 相差懸殊,是因為其輸送過程的性質或通過的介質大不相同。水在植物體內的輸送大體上可分為蒸騰、水分體內運輸、根系對水分的吸收等環節。
蒸騰作用水分通過植物表皮向大氣擴散的過程稱為蒸騰作用 。根據擴散的通路又可分為氣孔蒸騰 、角質層蒸騰、皮孔蒸騰 。其中氣孔蒸騰在氣孔開放時可占總蒸騰量的80~90%,但氣孔的開張度隨植株內外環境而變化。夜間或夏天中午炎熱乾旱時氣孔關閉,阻力增加,蒸騰速率很低。
植物體內水分運輸 蒸騰作用 草本植物 可超過數倍。大量蒸騰失水的補足,需有良好的通道。質外體中的細胞壁由親水的纖維素、半纖維素和果膠質 組成,其中可充滿水分,又無不透水的胞膜隔斷,水通過的阻力很小。在較長距離的水分運輸中,木質部 中的導管 是主要的通道。導管由一系列中空的已死細胞組成。水以連續的水絲形式集體流動,因而阻力很小。水絲從根的中柱鞘開始,直到葉肉細胞,與整個質外體中的水形成一個連續體。高大的樹木上層的葉片與根部之間的高度差可達百米。因為大氣壓只相當於10.34米水柱的壓力,蒸騰拉力 如何能將土壤中的水牽引到如此高的樹冠 ,而樹幹中的水絲不斷,在理論上曾經是個困難問題。英國H·H·狄克遜於1895年提出了內聚力 學說,他證明:液態水的分子間有很強的內聚力。如果密閉於管道中的水柱中沒有氣泡或空隙,就可以象固體棒那樣抵抗高達相當於300個大氣壓 的拉力而不斷。導管內有產生氣泡而使水絲中斷的機會,但導管細而多,組成導管的細胞或管胞之間有胞壁,從而避免了因一個小氣泡出現而使整個水絲中斷或氣泡向上下延伸的危險。局限在少數導管內的小氣泡可在夜間水分供應緩和時消失,從而使水絲恢復。導管的壁上有各種形式的加厚結構,可抵抗蒸騰拉力產生的負壓。但在蒸騰需求高時,負壓仍可使導管收縮、截面積變小。樹幹直徑每天因此表現明顯的變化,可以用樹徑記錄儀 來測量並記錄。因為通量與管徑的四次方成正比,導管直徑的收縮顯著增加水的運輸阻力。
白天根系吸水速率低於蒸騰失水速率時,植物體內水勢 下降,除導管體積縮小以外,整個質外體和與質外體接觸的共質體 ,也都交出一些水分。有些植物果實中的水也流出,進入蒸騰流 。夜間蒸騰降 低後,這些部分的水量又逐漸得到補充。因而可把這些部分看作水庫 。但除大樹和少數植物以外,庫容與一天的蒸騰量相比都不大。
根系對水分的吸收 根系對水分的吸收 土壤 水分相接觸。當葉片蒸騰失水時,水勢比土壤水那一端的水勢顯著降低,在植株體內形成水勢陡度,水分自土壤經根毛表面向根內流動。這種水分吸收順著水勢陡度發生,根系不起推動作用,因此稱為被動吸水 。當地上部不蒸騰,也沒有蒸騰所造成的低水勢來牽引蒸騰流時,根系也能吸收土壤中的水分,並推動水經木質部流向地上部。這種吸水過程稱為主動吸水 ,所產生的壓力稱為根壓 。在夜間和清晨空氣濕潤時,某些植物(特別是它們的幼苗和葉尖)分泌出水滴,稱為吐水;在切斷植物的莖時,連根部分的切端上木質部處形成水珠,稱為傷流。這兩種現象都是根壓造成的。根壓一般不超過0.2MPa,遠較蒸騰造成的拉力小,因此在植物吸水總量中貢獻很小,也不足以將水運到離地幾十米的樹冠上去。主動吸水要由呼吸作用提供能量。呼吸推動離子的主動吸收,使根細胞 中離子濃度高於土壤溶液之後,根細胞的水勢因而下降,吸引土壤溶液中的水進入根細胞。所以這裡水仍然是順著水勢差而流動的。水進入根系後,主要沿著運輸阻力較小的質外體運動,但根的內皮層有不透水的凱氏帶將質外體的通道阻斷,水只能通過屬於共質體的細胞質朝木質部方向移動。穿越胞膜時經過的阻力特別大。
除根以外,植物地上部也能吸收水分。在少雨而多霧和露的特殊地區,如智利 、秘魯 安第斯山脈海拔1000米高度之處的植物,水的主要來源是通過葉片 吸收的。
水分脅迫 水分脅迫 水分平衡 。前者大於後者時,植物含水量下降,水勢和膨壓也相應降低。超過一定限度時,植物的正常生理過程就會受到干擾,甚至使植物遭受損傷,這種水分虧缺稱為水分脅迫 或水分逆境。土壤水分過多也對植物造成傷害,也是脅迫,但那是由於土壤漬水阻斷根系的氧氣供應,妨礙有氧呼吸而造成的。植物各項生理功能對水分脅迫的敏感性差別很大。生長(特別是細胞膨大階段)最為敏感。溫室中生長的,正在伸展的玉米 與向日葵 葉片水勢只要比供水充分的葉片低0.2~0.3MPa就足以使生長明顯減緩。細胞壁合成、細胞分裂、蛋白質合成和硝酸還原酶的活力等也對水分脅迫敏感。水分脅迫還引起脫落酸合成量大大增加;乙烯釋放量增多;氣孔關閉,光合作用減弱;以及花、果、葉脫落。中生植物萎蔫時,體內可溶性糖和胺基酸 特別是脯氨酸 含量明顯增加。同時一些水解酶 從相應的區隔中釋放出來,因而產生了破壞作用。更嚴重的水分脅迫最終將導致生物膜系統嚴重損壞,造成植物死亡。植物對水分脅迫有多種抗禦的功能,就其與脅迫的關係可以分為三大類:①逃避,例如沙漠中的短命植物,在一次降雨之後,短時期(一個月)內就完成從種子萌發到開花結籽的整個周期。植物實際上不直接經受水分脅迫;②迴避,植物雖經受水分脅迫,但以某些回響防止了體內不利影響的發生。例如乾旱時氣孔 關閉,防止了水分的散失和體內水勢的下降;根冠比增高使供應單位葉面積的根吸收表面積增加,從而改變供求比等;③忍耐,變水型旱生植物能忍受強度脫水,直到氣乾狀態仍不死亡;再獲雨水時能很快恢復生命活動,也稱為復甦植物 。恆水植物中北美南部沙漠區的Larrea tridentata,旱季中老葉和小枝脫落,只留下長成的葉和芽,含水量降到乾重的50%也不引起嚴重損傷,雨後仍能重新生長。

 植物水分生理
植物水分生理 植物生理學
植物生理學 水分子
水分子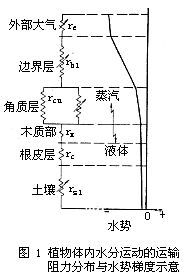 植物水分生理
植物水分生理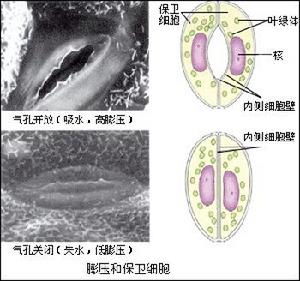 蒸騰作用
蒸騰作用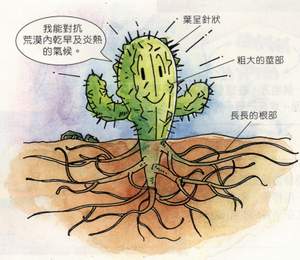 根系對水分的吸收
根系對水分的吸收 水分脅迫
水分脅迫