
概念
順反子(cistron):即結構基因,為決定一條多肽鏈合成的功能單位。1955年,美國分子生物學家本澤(Benzer)通過對大腸桿菌的噬菌體T4的RII區基因的深入研究,揭示了基因內部的精細結構。提出了基因的順反子(Cistron)概念。他發現,在一個基因內部,可以發生若干不同位點的突變,倘若在一個基因內部發生兩個以上位點的突變,其順式和反式結構的表型效應是不同的。
1957年,美國分子生物學家西莫爾·本澤爾(seymourBenzer,1921--)以T4噬菌體為材料,在DNA分子水平上研究基因內部的精細結構,提出了順反子(cistron)、突變子(muton)和重組子(recon)的概念。順反子是一個遺傳功能單位,一個順反子決定一條多肽鏈,這就使以前一個基因一種酶的假說發展為一個基因一種多肽的假說。能產生一種多肽的是一個順反子,順反子也就是基因的同義詞。順反子可以包含一系列突變單位──突變子。突變子是DNA中構成基因的一個或若干個核苷酸。由於基因內的各個突變子之間有一定距離,所以彼此間能發生重組,這樣,基因就有了第三個內涵──“重組子”。重組子代表一個空間單位,它有起點和終點,可以是若干個密碼子的重組,也可以是單個核苷酸的互換。如果是後者,重組子也就是突變子。順反子概念把基因具體化為DNA分子的一段序列,它負責傳遞遺傳信息,是決定一條多肽鏈的完整的功能單位;但它又是可分的,組成順反子的核苷酸可以獨自發生突變或重組,而且基因與基因之間還有相互作用。基因排列位置的不同,會產生不同的效應。
分類
單順反子
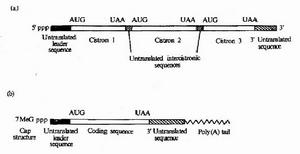 真核生物的為單順反子
真核生物的為單順反子單順反子(monocistron):真核基因轉錄產物為單順反子,即一個基因編碼一條多肽鏈或RNA鏈,每個基因轉錄有各自的調節元件。
真核生物mRNA(細胞質中的)一般由5'端帽子結構、5'端不翻譯區、翻譯區(編碼區)、3'端不翻譯區和3'端聚腺苷酸尾巴構成。分子中除G構成帽子外,常含有其他修飾核苷酸,如A等。5'端帽子結構通常有3種類型,即:G(5')ppp(5')N;G(5')ppp(5')N和G(5')ppp(5')N。圖1b[真核生物mRNA結構示意圖b5'端帽子結構式,表示鹼基,表示鹼基是帽子的化學結構,N右邊的m代表核糖2'位羥基的甲基化。真核細胞線粒體中的mRNA無帽子結構。一般認為帽子的功能與翻譯的啟動有關。許多真核生物mRNA(如珠蛋白mRNA)除去帽子後翻譯效率大大降低。5'端不翻譯區,也叫前導順序。不同的真核mRNA的前導順序長度不同,有的只有10個核苷酸,有的則有200個核苷酸。與原核mRNA相似,真核mRNA5'端不翻譯區中常有一段順序與核糖體小亞基上的18SrRNA的3'端的一段順序互補並結合,這種結合與真核mRNA的翻譯啟動有關。
翻譯區(編碼區)使用的密碼子除線粒體(如人、牛和酵母線粒體)外與原核生物mRNA是一樣的。真核生物mRNA的起始密碼子都是AUG。真核和原核生物mRNA使用的密碼子也都有“簡併現象”,即幾種不同的密碼子翻譯出同一種胺基酸,但不同的mRNA中簡併密碼子的利用率是不同的,真核與原核生物之間的差別就更大。mRNA的終止密碼子有3個(UAG、UGA和UAA),其功能是停止翻譯,一般只用一個終止密碼子就能使翻譯停止。有的mRNA有2個連續的終止密碼子(見)。3'端不翻譯區的長短在不同的mRNA上有所不同,β珠蛋白mRNA只有39個核苷酸,而卵白蛋白mRNA則有637個核苷酸。真核生物mRNA3'端不翻譯區常有AAUAA(A)或AUUUA(A)等順序,它們和識別多聚A聚合酶及裝配多聚A尾巴有關。除個別組蛋白mRNA外,真核生物mRNA3'端均有多聚A尾巴3'端多聚A尾巴的長度隨來源不同而不同,且隨mRNA的老化而變短,通常有20~200個A多聚A與mRNA穩定性及mRNA從細胞核轉到細胞漿中有關。
多順反子
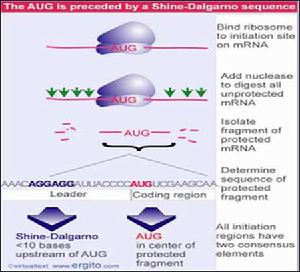 多順販子
多順販子在原核細胞中,通常是幾種不同的mRNA連在一起,相互之問由一段短的不編碼蛋白質的間隔序列所隔開,這種mRNA叫做多順反子mRNA。順反子的概念來自遺傳學中的順反重組試驗,是確定交換片段究竟在一個基因內還是屬於兩個基因的試驗,簡言之,一個順反子就是一個基因,多順反子就是多個基因。真核生物中也有多順反子,比如C.elegans共有13500個基因,約25%的是多順反子(polycistronicmRNA)。
mRNA存在於原核和真核生物的細胞質及真核細胞的某些細胞器(如和)中。RNA病毒和RNA噬菌體中的RNA既是遺傳信息的載體又具有mRNA的功能。生物體mRNA種類的多少與生物進化水平有關,高等生物所含的遺傳信息多,mRNA的種類也多。生物體內某種mRNA的含量根據需要而有不同,如5齡蠶後部絲腺體的主要任務是快速合成大量絲心蛋白,因而編碼絲心蛋白的mRNA含量特別多。有些細菌需要不斷適應外部環境,其體內編碼某些誘導酶的mRNA的含量也較多。
原核和真核生物mRNA有不同的特點:①原核生物mRNA常以多順反子(見)的形式存在,即一條mRNA鏈編碼幾種功能相關聯的蛋白質。真核生物mRNA一般以單順反子的形式存在,即一種mRNA只編碼一種蛋白質。②原核生物mRNA的轉錄與翻譯一般是偶聯的,即轉錄尚未完畢,蛋白質的轉譯合成就已開始。真核生物轉錄的mRNA前體則需經後加工,加工為成熟的mRNA與蛋白質結合生成信息體後才開始,工作信息體中蛋白質與RNA之比約為3。③原核生物mRNA半壽期很短,一般為幾分鐘,最長只有數小時(RNA噬菌體中的RNA除外)。真核生物mRNA的半壽期較長,如胚胎中的mRNA可達數日。④原核與真核生物mRNA的結構特點也不同。
一級結構與功能的關係:原核生物mRNA一般5'端有一段不翻譯區,稱前導順序,3'端有一段不翻譯區,中間是蛋白質的編碼區,一般編碼幾種蛋白質。如大腸桿菌乳糖操縱子mRNA編碼3條多肽鏈;色氨酸操縱子mRNA編碼5條多肽鏈。也有單順反子形式的細菌mRNA,如大腸桿菌脂蛋白mRNA。原核生物mRNA分子中一般沒有修飾核苷酸,也沒有5'端帽子結構和3'端聚腺苷酸尾巴。在原核生物mRNA的起始密碼子(AUG)附近(5'方向上游)的一小段長短不等的順序,含有較多的嘌呤核苷酸,被稱為SD順序。它能和核糖體小亞基上的16SrRNA的3'端富含嘧啶核苷酸的區域配對結合,有助於帶有甲醯甲硫氨酸的起始tRNA識別mRNA上的起始密碼(AUG),使肽鏈合成從此開始。這段順序是1974年由J.夏因和L.達爾加諾發現的,所以稱為SD順序,也稱核糖體結合部位。原核生物mRNA的編碼區一般編碼幾種功能上相關聯的蛋白質,兩種蛋白質的編碼區之間常有一小段不翻譯的順序,叫做間隔區。有的噬菌體RNA中2個相鄰的順反子共用一段相同的編碼順序,例如,M噬菌體RNA中的溶菌蛋白編碼區共225個核苷酸中有189個核苷酸是由相鄰兩個蛋白質共用的。原核mRNA與真核mRNA一樣使用同一套三聯體密碼子(真核生物線粒體mRNA有例外)。原核生物合成胺基酸的操縱子mRNA的5'端前導順序上有一段順序稱作弱化子。弱化子具有兩種可以互變的構象,其中一種構象是轉錄終止的信號,能使轉錄中止(或衰減)。衰減調節是原核生物合成胺基酸的調控方式之一。
套用前景
1、根據已發表的相關序列,分別設計引物,通過PCR技術克隆了一系列構建水稻質體多順反子定點整合表達載體所需的元件:質體核糖體(16S)RNA操縱元啟動子(Prrn)、質體psbA基因3′端終止子(psbA3′)、氨基糖苷3′-腺苷醯基轉移酶基因(aadA)、水稻質體基因組高頻同源重組片段(psbC/trnG,大小3362bp),命名為crDNA、甘露聚糖酶基因(man)、綠螢光蛋白基因(gfp)。構建了水稻質體多順反子定點整合表達載體pLM21(-psbC-Prrn-RBS-man-RBS-gfp-RBS-aadA-psbA3′-trnG-)。將該載體用基因槍轟擊菸草葉片5槍,用添加了壯觀黴素的選擇分化培養基篩選。結果獲得質體轉基因菸草4株。用PCR、雷射掃描和Westernblot等方法檢測都證實man、gfp、aadA三個基因且均得到表達,用RFLP證實表達盒整合到菸草質體基因組中。
2、利用腦心肌炎病毒(EMCV)及脊髓灰質炎(polio)病毒核心糖體進入位點(IRES),連線mIL-12p40及p35cDNAs和篩選基因新黴素磷酸轉移酶(NeoR),克隆至逆轉錄病毒載體pGCEN中,使三個基因同時受逆轉錄病毒載體5′端LTR啟動子控制,轉錄至同一mRNA轉錄本上,通過不同機制翻譯成蛋白質,從而構建成多順反子逆轉錄病毒載體,即pGCEN/mIL-12。在LipofectAMINE介導下將pGCEN/mIL-12轉染包裝細胞PA317,G418篩選,直至出現陽性克隆,挑取抗性克隆,擴大培養,收集上清,用小鼠成纖維細胞NIH3T3測定病毒滴度。然後用重組轉錄病毒感染小鼠肝癌細胞MM45T.Li.G418篩選,直至出現抗性克隆,擴大培養,對陽性克隆進行鑑定。結果是由PA317包裝細胞產生的重組逆轉錄病毒的滴度為5×105CFU/ml,將其感染小鼠肝癌細胞MM45T.Li,後經PCR及Southernblot證明,外源基因已整合至小鼠肝癌細胞基因組中,RT-PCR及Northernblot分析外源基因在mRNA水平上的表達,並證實mIL-12p40及p35cDNA和NeoR基因轉錄在同一mRNA上。ELISA顯示mIL-12的表達量48h為10ng/106細胞。並且M45/mIL-12培養上清能刺激人外周血單個核細胞增殖及誘導小鼠脾細胞IFN-γ的產生(160U/ml)。
3、目前一種新的基因治療策略,即將不同耐藥譜基因導入正常骨髓造血幹細胞之獲得耐藥表型,加大造血細胞與腫瘤細胞克隆之間對化療耐受性的差別,增加骨髓對化療的耐受性,以便能施行大劑量化療、多次重複給藥使之最大限度清除腫瘤細胞,發揮化療效,為保護和及時恢復造血功能提供了可行性。
本研究項目中造血幹細胞採用的是臍血幹細胞[wangjishi,etal.LeukemiaReaearch.2002,26(3)281-288;實驗生物學報2001,34(3)227-233]\外周血CD34+細胞[wangjishi,FangQinetaL.AxtaPharmacologicaSinica.2001,22(10):949-955]和小鼠骨髓細胞,轉染靶細胞顯示對化療藥物的抗性增加。有效表達,增加了細胞對亞硝基類藥物(如BCNU)的耐愛性,證實該基因具有明顯的細胞生物學效應。同進通過體外誘導突變技術獲得了突變型MGMT,並成功構建到逆轉錄病毒載體上。本研究為同內首次以RT-PCR方法從人肝組織克隆了06-甲基鳥嘌呤-DNA-甲基轉移酶(MGMT)基因,與人類基因文庫MGMT基因同源性比為100%,將該基因導入靶細胞後顯示有效表達,增加了細胞對亞硝基類藥物(如BCNU)的耐受性,證實該基因具有明顯的細胞生物學效應。同進通過體外誘導突變技術獲得了突變型MGMT,並成功構建到逆轉錄病毒載體上。
本研究課題構建了國內外尚未報導的含ALDH1、ALDH3基因與IRES-MDRI基因的雙順反子逆病毒表達載體。同時證實了ALDH1、ALDH3均有抗活性環磷醯胺作用。在多順反子逆病毒表達載體GLNA-MGMT-NEOR-IRES-MDRI中,利用NEOR基因和MDRI的雙重作用建立了雙標記系統。研究中涉及到了DNA序列分析,桌球效應,RT-PCR,PCR,NORTHERNBLOT,WESTERNBLOT,FACS,MTT、建立高滴度產病毒細胞體系和安全高效的基因轉移系統。
