
分布
分布較廣;澳洲、歐洲、日本、北美、南美和南非都有報導。
寄主植物
自然寄主範圍很窄,主要是藜科植物,很多都是產生褪綠斑駁,但是歐洲也發現很多果樹和葡萄也感染藜草花葉病毒。有實驗證明葡萄藤的分離株可經實驗方法侵染其它科植物(Bercks&Querfurth,1969)。
Chenopodium spp:葉片褪綠斑駁
Vitis spp.,Prunusdomestica:幾乎不侵染
Atriplex suberecta:矮化,葉片變形
實驗寄主範圍和症狀
莧色藜(ChenoPodbo amaranticoI):機械接種的植株先出現褪綠局斑(圖3) 然後出現黃色斑點和星狀病斑(圖1、2)。經種子感染的植株出現斑駁和斑點症狀,也可能不表現任何症狀。單用水或緩衝液潤濕的手指抹擦葉片,不可能引起病害症狀(Dias &Whterwort,1967;Engelbrecht &van Regenmortel,1968)
昆諾藜和牆生藜的症狀與莧色藜的症狀相似。
甜菜(Beta vulgaris)非常輕微的系統花葉。
繁殖寄主
莧色黎、昆諾藜或牆生藜都適於保存病毒。
菠菜(SPinacia oleracea)是純化病毒的良好材料。
鑑定寄主
莧色藜和昆諾藜都出現局斑,但是否適於作為鑑定寄主,還未曾試驗。
危害情況
不少藜屬植物產生褪綠斑駁,其中許多種類可作為其他植物病毒的實驗測定寄主。實驗植物,如莧色藜(ChenoPodbo amaranticoI)、昆諾藜(C.quinoa)和牆生藜(C.murale),都可帶這種病毒,但經種子傳播無症狀表現。
經濟重要性
SoMV在雜草的經濟影響較小。栽培作物和觀賞植物很少受到病毒侵染,經濟作物如被侵染,症狀一般為和性花葉或無症狀。
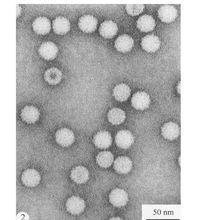
形態特徵
質粒結構
RNA病毒,質粒等軸對稱,無包膜,直徑 2 6nm。葉子汁液含有很多病毒粒子。有些分離物病毒粒子在pH4的條件下比pH7更穩定。經磷鎢酸或醋酸鈾染色的質粒的電鏡照片分辨不出任何形態亞基(圖4),
只有一種沉降組分。高度稀釋下的沉降係數s(20,w):104±2S
 藜草花葉病毒
藜草花葉病毒分子量:6.3 ± 0.4 × 106(Kado&Black,1968)。
擴散係數:(D20/10-7厘米2/秒):l.2 9(根據 Kado計算,未發表);1.15(Paul&
Huth,1970)。
等電點:PH4.4。
部分比容(計算)0.70 ±0.05厘米3/克。
260毫微米吸收值(毫克/毫升,1厘米光程):4.9。
260/280比值(未校正光散射):1.49;230/260比值;1.67。
質粒組成
RNA:分予量約1.3 ×106,約占質粒重的 2 0%,可能是單鏈。核苷酸的克分子百分比:G 26.0,A 23.2,C 27.6,U 23.2。
蛋白質:
質粒僅含一種蛋白質亞基。C末端胺基酸是賴氨酸,N末端封閉。胺基酸組成(克分子%): 丙氨酸8.5;精氨酸4.6;天冬氨酸9.1;半胱氨酸1.1;谷氨酸6.8;甘氨酸9.1;組氨酸1.7;異亮氨酸5.1; 亮氨酸6.8;賴氨酸6.8;甲硫氨酸2.8;苯丙氨酸2.3;脯氨酸6.8;絲氨酸8.0;蘇氨酸7.4;色氨酸1.7;酪氨酸4.0;纈氨酸7.4(Kado,1967)根據胺基酸分析提出,蛋白質亞基的分子量為 l.92× 104(Kado,1967)。根據中性十二烷基硫酸鈉/聚丙烯酸胺凝膠電泳指出,蛋白質亞基的分子量可能等於3.1× 104(Hill,1971)。
汁液穩定性
昆諾藜汁液中,病毒熱鈍化點(10分鐘)約90 ℃,稀釋限點約10-7。在23℃下保持侵染性約60天。在冰凍條件下,侵染性可長期保持。
生物學
株系
所有分離株的血清學反應都一致。這些分離株包括:KirkPatriok等(1965)及 Bancroft和 Tolin(1967)的蘋果隱症病毒;Holling和 Stone(1967)的香石竹 698號分離株;Silye等(1958)的藜花葉病毒;Kado(1966;1967)的藜星斑病毒;Teakle(1968)的魚草(ChenoPodlum trigonon)病毒;葡萄藤的分離株(Bercks&Querfurth,1969)。除了葡萄藤分離株外,其它所有的分離株都很相似,因為葡萄藤分離株可經實驗方法感染莧科、葫蘆科、豆科和茄科植物
血清學
病毒的免疫原性很強。緩衝液含1%瓊脂或瓊脂糖凝膠的雙擴散試驗,試管中的沉澱試驗,1%瓊脂或瓊脂糖凝膠的免疫電泳試驗,都可以用。通常在凝膠雙擴散試驗和免疫電泳試驗中,形成一條沉澱帶。試管沉澱試驗中,沉澱呈顆粒狀。
親緣關係
未檢出該病毒與下列病毒之間有血清關係:苜蓿花葉病毒、南芥草花葉病毒、菜豆莢斑駁病毒、蠶豆斑駁病毒、豇豆花葉病毒、黃瓜花葉病毒、黃瓜花葉病毒Y毒株、葡萄扇葉病毒、桃黃芽花葉病毒、南方菜豆花葉病毒,南瓜花葉病毒、菸草壞死病毒、番茄斑萎病毒、蕪青黃花葉病毒和野黃瓜花葉病毒。
病毒與細胞和組織的關係
所有的組織都能感染病毒。細胞質內的病毒呈膜包裹的晶狀排列,引起"假核仁"一緻密、環形、粒狀的小區(Milne,1967)。
細胞病理學
病毒顆粒在葉肉細胞和木質部都有發現,但在韌皮部沒有發現病毒粒體。在單個的細胞中病毒存在於細胞質和液泡中。受侵染細胞的內含體在細胞質中呈晶體狀
純化
每公斤接種的葉組織可得病毒200~500毫克。
下列兩種方法均可採用:
1.Kado(1967)的方法:用 pH7.5的 0.05M二乙胺四乙酸鈉研磨冰凍組織。過濾後,加入0.5倍體積的冷正丁醇,攪拌,經離心使乳濁液澄清。吸出上層的正丁醇,用玻璃棉過濾液相。高低速離心5次,使病毒濃縮,將病毒沉澱再懸浮在PH7.0的0.1M磷酸緩衝液中。
2.Steer(1956)的丁醇一氯仿法。
傳播途徑
介體傳播
據報導,豌豆潛葉蠅(Liriomyza langei)、甜菜葉蟬(Circulifer tenellus)一種蝽(Halticus citri)和桃蚜(Myzus Persicae)都可能是介體(Bennet& Costa,1961)。然而Dias和Waterworth)沒能證實桃蚜傳毒。
種子傳播
病株子代幼苗的感染率高達83%,感染率的高低取決於種子的取樣(Bennett&Costa,1961;Dias&waterworth,1967 ;Kado,1967)。通過種子而感病的植株很少出現症狀,甚至毫無症狀。
菟絲子傳播
加州菟絲子(cuscuta californica)不傳毒,它顯然對感染是免疫的(Bennett&Costa,1961)。
檢疫與防治
檢測方法
因為SoMV經常是潛隱或症狀較輕,檢測時要轉接到無SoMV的指示植物如莧色藜、昆諾藜,然後進行血清學檢測,或者是以血清學方法在初始寄主上檢測。檢測系統侵染的寄主如Chenopodium spp,因為病毒含量很高,較易檢測,而在另外一些寄主上,因病毒含量很低且移動很慢,即使用DAS-ELISA檢測也較為困難(Cardin et al., 1995)。
防治
很多時候,因為SoMV只產生很輕的症狀,發作也是零星的發作,因此認為對其的控制不是很重要。它能引起的問題是污染莧科指示植物的種子,使其無症帶毒,這會給以後的研究帶來不便。
無化學或其他控制介體的方法。
注
藜屬植物的種子可能攜帶藜花葉病毒,這一事實表明,當這些品種的植物作為其它病毒的寄主時,會出現污染的問題,尤其因為感病的子代幼苗可能不出現什麼症狀。所以,從表面上看似乎不帶藜花葉病毒的植株上收集種子時,務須謹慎。
