
發現
支原質體是在1898年發現的,是一種簡單的原核細胞。其大小介於細菌和病毒之間。結構也比較簡單,多數成球形,沒有細胞壁,只有三層結構的細胞膜,故具有較大的可變性。支原質體可以在特殊的培養基上接種生長,用此法配合臨床進行診斷。與泌尿生殖道感染有關的主要是分解尿素支原質體和人型支原質體兩種,約有20-30%的非淋菌性尿道炎的病人,是由以上兩種支原質體引起的,是非淋菌性尿道炎及宮頸炎的第二大致病菌。在成年人的泌尿生殖道中分解尿素支原質體和人型支原質體感染率主要與性活動有關,也就是說,與性交次數的多少、性交對象的數量有關,不管男女兩性都是如此。據統計女性的支原質體感染率更高些,說明女性的生殖道比男性生殖道更易生長支原質體。另外,分解尿素支原質體的感染率要比人型支原質體的感染率為高。性狀
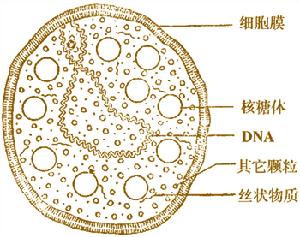 形態結構
形態結構支原質體的大小為0.2~0.3um,可通過濾菌器,常給細胞培養工作帶來污染的麻煩。無細胞壁,不能維持固定的形態而呈現多形性。革蘭氏染色不易著色,故常用Giemsa染色法將其染成淡紫色。細胞膜中膽固醇含量較多,約占36%,對保持細胞膜的完整性具有一定作用。凡能作用於膽固醇的物質(如二性黴素B、皂素等)均可引起支原質體膜的破壞而使支原質體死亡。
支原質體基因組為一環狀以雙鏈DNA,分子量小(僅有大腸桿菌的五分之一),合成與代謝很有限。
肺炎支原質體的一端有一種特殊的末端結構(terminal structure),能使支原質體粘附於呼吸道黏膜上皮細胞表面,與致病性有關。
培養特性
營養要求比一般細菌高,除基礎營養物質外還需加入10~20%人或動物血清以提供支原質體所需的膽固醇。最適pH7.8~8.0之間,低於7.0則死亡,但解脲脲原體最適pH6.0~6.5。
大多數皆性厭氧,有些菌株在初分離時加入5%CO2生長更好。生長緩慢,在瓊脂含量較少的固體培養基上孵育2~3天出現典型的“荷包蛋樣”菌落:圓形(直徑10~16um),核心部分較厚,向下長入培養基,周邊為一層薄的透明顆粒區。此外,支原質體還能在雞胚絨毛尿囊膜或培養細胞中生長。
繁殖方式多樣,主要為二分裂繁殖,還有斷裂、分枝、出芽等方式,蓋因缺乏細胞壁造成分裂時二個子細胞大小均所致。同時,支原質體分裂和其DNA複製不同步,可形成多核長絲體。
支原質體分化
生化反應與分型
一般能分解葡萄糖的支原質體則不能利用精氨酸,能利用精氨酸的則不能分解葡萄糖,據此可將支原質體分為兩類。解脲脲原體不能利用葡萄糖或精氨酸,但可利用尿素作能源。
各種支原質體都有特異的表面抗原結構,很少有交叉反應,具有型特異性。套用生長抑制試驗(Growth inhibition test,GIT)、代謝抑制試驗(Metabolic inhibition test,MIT)等可鑑定支原抗原,進行分型。
抵抗力
支原質體對熱的抵抗力與細菌相似。對環境滲透壓敏感,滲透壓的突變可致細胞破裂。對重金屬鹽、石炭酸、來蘇爾和一些表面活性劑較細菌敏感,但對醋酸鉈、結晶紫和亞銻酸鹽的抵抗力比細菌大。對影響壁合成的抗生素如青黴素不敏感,但紅黴素、四環素、鏈黴素及氯黴素等作用於支原質體核蛋白體的抗生素,可抑制或影響蛋白質合成,有殺滅支原質體的作用。
致病性與免疫性
支原質體不侵入機體組織與血液,而是在呼吸道或泌尿生殖道上皮細胞粘附並定居後,通過不同機制引起細胞損傷,如獲取細胞膜上的脂質與膽固醇造成膜的損傷,釋放神經(外)毒素、磷酸酶及過氧化氫等。
巨噬細胞、lgG 及 lgM 對支原質體均有一定的殺傷作用。呼吸道黏膜產生的SlgA抗體已證明有阻止支原質體吸附的作用。在兒童中,致敏淋巴細胞可增強機體對肺炎支原質體的抵抗力。
感染
致病支原質體中,肺炎支原質體起肺炎,人型支原質體、解脲支原質體和生殖器支原質體主要泌尿生殖道感染。支原質體肺炎又稱原發性非典型肺炎,支原質體肺炎全年均可發病,以冬季多見,可有小流行。支原質體腦炎是學齡前兒童及青年人常見的一種肺炎,支原質體肺炎主要通過飛沫傳播,潛伏期較長,可達2~3周。支原質體肺炎雖然病程較長,肺部病變較重,炎症吸收較慢,但絕大多數預後都是良好的,合併症亦少。生殖器支原質體感染是近年新明確的一種性接觸傳播疾病。成人主要通過性接觸傳播,新生兒則由母親生殖道分娩時感染。成人男性的感染部位在尿道黏膜,女性感染部位在宮頸。新生兒主要引起結膜炎和肺炎。診斷
 診斷
診斷測定支原質體抗體的血清學試驗方法中,有支原質體特異性血清學檢測和非特異性血清學檢測:支原質體特異性血清學檢測方法中,最常用的是補體結合試驗,另有間接免疫螢光染色檢查法、生長抑制試驗、代謝抑制試驗、間接血凝試驗、酶免疫法和酶聯免疫吸附試驗(ELISA)等。支原質體的非特異血清學方法有肺炎支原質體冷凝集試驗與MG鏈球菌凝集試驗,對支原質體肺炎能起輔助診斷的作用。檢測特異性抗體IgG的方法尚不能達到早期快速診斷的目的,抗原的檢測為今後研究的發展方向。目前已有用酶聯免疫吸附試驗、螢光標記抗體、肺炎支原質體膜蛋白單克隆抗體和反向間接血凝法直接檢測分泌物和體液中支原質體抗原的報導,具有很高的特異度和靈敏度。人體感染肺炎支原質體後,能產生特異性IgM和IgG類抗體。IgM類抗體出現早,一般在感染後1周出現,3~4周達高峰,以後逐漸降低。由於肺炎支原質體感染的潛伏期為2~3周,當患者出現症狀而就診時,IgM抗體已達到相當高的水平,因此IgM抗體陽性可作為急性期感染的診斷指標。如IgM抗體陰性,則不能否定肺炎支原質體感染,需檢測IgG抗體。IgG較IgM出現晚,需動態觀察,如顯著升高提示近期感染,顯著降低說明處於感染後期。
培養
 固體培養基上的菌落特徵
固體培養基上的菌落特徵支原質體污染的來源包括工作環境的污染、操作者本身的污染(一些支原質體在人體是正常菌群)、培養基的污染、污染支原質體的細胞造成的交叉污染、實驗器材帶來的污染和用來製備細胞的原始組織或器官的污染。
組織細胞培養工作中,主要從以下幾個方面來預防支原質體的污染:控制環境污染;嚴格實驗操作;細胞培養基和器材要保證無菌;在細胞培養基中加入適量的抗生素。支原質體污染細胞後,特別是重要的細胞株,有必要清除支原質體,常用方法有抗生素處理、抗血清處理、抗生素加抗血清和補體聯合處理。支原質體最突出的結構特徵是沒有細胞壁,一般來講,對作用於細胞壁生物合成的抗生素,如 -內醯胺類、萬古黴素等完全不敏感;對多粘菌素(polymycin)、利福平、磺胺藥物普遍耐藥。對支原質體最有抑制活性及常用於支原質體感染治療的抗生素是四環素類、大環內酯類及一些氟喹諾酮;其他類抗生素如氨基糖苷類、氯黴素對支原質體有較小抑制作用,所以常不用來作為支原質體感染的化學治療劑。
支原質體有時被稱為類菌質體。無細胞壁,多呈不規則球狀、長絲狀,可分枝,營寄生共生或腐生。一般侵害對象為動植物,可造成多種疾病。
支原質體(Mycoplasmal)是目前所能發現的能在無生命培基中生長繁殖的最小的微生物。 支原質體體形多樣,基本為球形,亦可呈球桿狀或絲狀,其菌落呈針尖大小,故又稱之為微小支原質體。支原質體特點是無細胞壁及前體,細胞器極少。DNA的G C含量低,菌體內具有非常小的染色體組,其分子量約為45×108,菌體細胞大小約為0.2-0.3μm,很少超過1.0μm。由三層蛋白質和脂質組成的膜樣結構以及一層類似毛髮結構組成。支原質體由二分裂繁殖,形態多樣。支原質體用普通染色法不易著色,用姬姆薩染色很淺,革蘭氏染色為陰性。支原質體可在雞胚絨毛尿囊膜上或細胞培養中生長。用培養基培養。營養要求比細菌高。由於它沒有細胞壁,因此對影響細胞壁合成的抗生素,如青黴素等不敏感,但紅黴素、四環素、卡那黴素、鏈黴素、氯黴素等作用於核蛋白體的抗生素,可抑制或影響支原質體的蛋白質合成,有殺傷支原質體作用,支原質體對熱抵抗力差,通常55℃經15分鐘處理可使之滅活。石碳酸,來蘇兒易將其殺死。在培養基中置入脲素並以硫酸錳作指示劑極易與其他支原質體作出鑑別。
它廣泛分布於自然界,有80餘種。與人類有關的支原質體有肺炎支原質體(M-pneumonie,Mp)、人型支原質體(M.humenis,MH)、解脲支原質體(Ureaplasma urealyticum, UU 分解尿素支原質體)和生殖器支原質體(M.genitalium,MG)等。
肺炎支原質體引起肺炎。現已從人類泌尿生殖道分離出來7種支原質體,其中分離率較高而與泌尿生殖道疾病有關,是解脲支原質體,其次是人型支原質體。人型支原質體(M.humenis,MH)、解脲支原質體(Ureaplasma urealyticum,UU)和生殖器支原質體(M.genitalium,MG)都會引起泌尿生殖道感染。
解脲支原質體(ureaplasma ure-alyticum)是一種原核微生物,呈球桿狀,大小為125-250毫微米,分子量4.5*108,高度多形性,沒有堅硬的細胞壁,能在無細菌的培養物中增殖,能產生尿素分解酶分解尿素。特異性抗體能抑制它生長,因其缺乏堅硬的細胞膜,對青黴素耐藥,對細胞膜有親和性,生長繁殖時需要類固醇物質。四環素、紅黴素、壯觀黴素對之有效。嬰兒或無性交接觸的女性生殖道內找不到解脲支原質體。而性生活越亂,這種支原質體陽性率也越高。Mc Donald 1982年報告587例急性尿道炎症狀病人中,209例(36%)中段尿中分離出解脲支原質體。
人型支原質體(mycoplasma hominis)支原質體對外界環境抵抗力弱,45℃15min即可被殺死。對肥皂、酒精、四環素、紅黴素敏感。
機理
 肺炎支原體
肺炎支原體在動物實驗發現,小鼠腹腔巨噬細胞可以殺滅支原質體,而中性粒細胞的作用不大。在體外,IgG1和IgG2抗體有調理作用。可加強巨噬細胞對支原質體的殺傷作用。
對比
肺炎支原質體
肺炎支原質體(M.Pneumonia)是人類支原質體肺炎的病原體。支原質體肺炎的病理改變以間質性肺炎為主,有時並發支氣管肺炎,稱為原發性非典型性肺炎。主要經飛沫傳染,潛伏期2~3周,發病率以青少年最高。臨床症狀較輕,甚至根本無症狀,若有也只是頭痛、咽痛、發熱、咳嗽等一般的呼吸道症狀,但也有個別死亡報導。一年四季均可發生,但多在秋冬時節。
肺炎支原質體的致病首先通過其頂端結構粘附在宿主細胞表面,並伸出微管插入胞內吸取營養、損傷細胞膜,繼而釋放出核酸酶、過氧化氫等代謝產生引起細胞的溶解、上皮細胞的腫脹與壞死。誘發機體產生的抗體也可能參與了上述病理損傷。呼吸道分泌的SlgA對再感染有一定防禦作用,但不夠牢固。
肺炎支原質體的診斷方法主要依靠分離培養和血清學試驗。標本可采可疑病人的痰或咽試子,接種於含血清或酵母浸膏的瓊脂培養基。5~10天后觀察有無直徑30~100um的圓形房頂樣菌落。多次傳代後可變為典型的“荷包蛋”樣菌落,並能吸附多種動物紅細胞和氣管上皮細胞、HeLa細胞等,且此類吸附可被特異性抗體所抑制。分離的支原質體經形態、溶血與生化反應作初步鑑定後需進一步用特異性抗血清作生長抑制試驗和代謝抑制試驗。用患者血清與支原質體脂質抗原作補體結合試驗,恢復期較急性期效價高4倍以上具有診斷價值。亦可用間接免疫螢光試驗、間接血凝ELISA檢測標本。另外,有1/3~3/4患者的血清可與人O型紅細胞在4℃時有非特異性凝集(稱為“冷凝集試驗”),37℃時消失,患病一周時達到高峰。此方法簡便,有助於診斷。
影響
 生殖道支原體
生殖道支原體支原質體經尿道感染後患者可出現尿道炎症狀,並可繼發慢性前列腺炎。在檢查前列腺液時,可見活潑、泳動的微生物群體。支原質體還繼續感染精道、精囊和睪丸,影響精子和精液的質量,引起不育。經觀察發現支原質體可通過下述環節引起不育症。
1 、干擾精子運動: 精子運動是健康精子的一項重要功能,是衡量精子能否受孕的重要指標,而且精子的運動必須有一定速度和頻率。支原質體感染精子後,常常附著在精子的頭部和尾部,使整個精子掛滿了大小不等的附著物,致使精子泳動無力,互相纏繞,導致不育。
2 、精子畸形率增加: 支原質體感染導致精子畸形率增加是造成不育的另一特徵。據臨床觀察,在這類不育患者中,精子畸形率有時可高達 80% 。
3 、破壞生精細胞: 睪丸的曲細精管中有大量生精細胞,這些生精細胞經過發育繁殖形成精子。當支原質體從尿道、前列腺等部位進入睪丸曲細精管後,會破壞生精細胞,使“生精工廠”產生偽劣產品,導致不育。
支原質體對女性生殖系統的影響
解脲支原質體可侵犯尿道、宮頸及前庭大腺,引起尿道炎、宮頸炎與前庭大腺炎;上行感染時,可引起子宮內膜炎、盆腔炎、輸卵管炎,尤其輸卵管炎多見。解脲支原質體感染造成的女性生殖器官病理性改變,是不孕不育的重要原因。國內外資料提示,不孕症夫婦的宮頸粘液、精液中解脲支原質體培養陽性率高達50%以上,由此可見,解脲支原質體感染與不孕症的發生有相關關係。解脲支原質體感染造成不良的另一個原因是流產,有人從流產的組織中檢查出解脲支原質體的陽性率高達40%以上。因此,對不明原因的流產,尤其是多次流產者,應考慮有解脲支原質體感染的可能。解脲支原質體感染造成的不完全梗阻的輸卵管炎性粘連,可使管腔狹窄,通而不暢,還是發生宮外孕的重要原因。
婦女妊娠後,由於孕激素的增加,抑制了細胞免疫,機體抵抗力下降,更易受到解脲支原質體的感染,解脲支原質體引起的圍產期感染已是現代產科面臨的新問題。解脲支原質體可以經胎盤垂直傳播或由孕婦下生殖道感染上行擴散,引起宮內感染,兩者均可導致流產、早產、胎兒宮內發育遲緩、低體重兒、胎膜早破,甚至造成胎死宮內等一系列不良後果。
