
形態發生運動
正文
在早期發育中,胚胎細胞的各種協調運動的總稱。通過這些運動,胚層建立了一定的空間關係,並為以後器官發生奠定基礎。形態發生運動包括原腸形成和器官原基形成。在動物胚胎髮育的囊胚期,甚至原腸早期,形成各種器官的物質都還沒有到達它們在成體中的部位(見胚胎預定命運圖)。通過複雜的運動和移位,原來位於表面的細胞進入內部,形成具有3個胚層的胚胎,外胚層留在外部,內胚層和中胚層移入內部,這一過程稱為原腸形成。在以後的發育中,3個胚層的細胞都將繼續發生一系列變化,以形成各種器官的原基,稱器官原基的形成(見器官發生)。原腸形成 1925年德國W.福格特套用活體染色法以後,人們對原腸形成運動才有了正確的了解。福格特不但繪製了兩棲類各胚層在囊胚期的位置,並且追蹤它們以後發育成什麼組織或器官。在此基礎上,經過方法上的改進,逐步了解了各種動物胚胎的形態發生運動。
原腸形成的過程因卵子卵黃含量的多寡而不同,含量少的,進程比較簡單,反之就比較複雜。
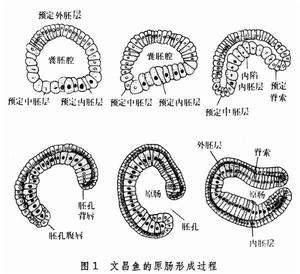
原索動物文昌魚的卵子,含卵黃較少,囊胚的構造是一層長柱形細胞包圍著一個腔──囊胚腔。文昌魚原腸胚的形成,是直接由一個單層的空腔變成雙層的結構(圖1)。首先是囊胚的植物極變為扁平,然後逐漸向內部陷入,這一種運動叫做內陷。內陷越來越深入,囊胚腔被擠得越來越小,最後,內層和外層相貼,囊胚腔完全消失,形成一個具有內外兩層細胞的杯形結構。內層包圍的腔是新形成的原腸腔,其與外界相通的開口稱胚孔。配合著內陷,外胚層也向下包圍生長──這一種運動叫做外包。由於外包和內陷是協調進行的,所以由單層變為雙層的胚胎不是半球形,而是大體上維持著原來大小的球形。外層就是外胚層包括將來形成表皮和神經系統的部分。內層包括脊索中胚層和內胚層,脊索沿前後方向伸長,占據背方中部的位置,中胚層位於兩側,它們共同形成原腸的頂部,而內胚層形成原腸的底部和側部。
 形態發生運動
形態發生運動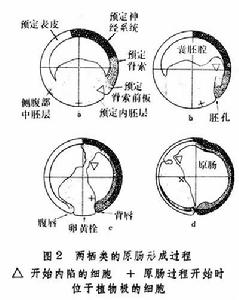 形態發生運動
形態發生運動原腸形成結束之後,有些物質,特別是形成軀幹後端和尾部體節的物質,還留在表面(見胚胎預定命運圖)。這些物質以後進入內部的運動甚至在背部的神經板形成之後還未完成。
R.E.凱勒認為,在無尾類爪蟾的囊胚預定脊索和中胚層細胞不是位於表面而是位於深層。在原腸形成開始,當內胚層沿背唇內卷的時候,位於它下面的中胚層一起捲入。爪蟾的情況是否也適用於其他兩棲類,尚待進一步驗證。
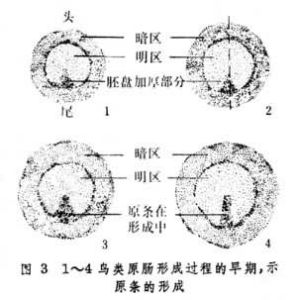
在卵黃含量豐富的爬行類和鳥類,內卷不再是原腸形成的一種方式。在鳥類,卵裂的結果在卵黃的頂端形成盤狀的囊胚,由上胚層和下胚層組成。所謂囊胚腔,是介於上胚層和下胚層之間的腔隙,從上面看,上胚層的中央部分,因為下面有囊胚腔襯托著,顯得比較明亮,稱為明區。所有的外胚層、中胚層和內胚層的細胞都來自上胚層,中胚層和內胚層細胞的移入局限在盤狀胚胎中線的一定部位。最初,上胚層的細胞向明區末端會聚,形成加厚的部份,然後向前延伸成條狀構造,即原條(圖3)。原條的當中有一條凹陷,稱為原溝。原溝底部的細胞呈瓶狀,其外端和表層相連,另一端向下伸出,將來形成中胚層和內胚層的細胞,就這樣以單個細胞的形式,在原溝的底部自上胚層向下移入(圖4)。當移入的細胞相互接觸時,又重新形成細胞層,中胚層和內胚層分別形成。原條伸展達到一定長度,其前端出現加厚部份,稱為原結,將來形成脊索的物質集中在這裡。原條加長,原結隨著向前移動,移入的細胞在上胚層之下形成“頭突”,逐漸伸長發育為脊索。在它附近經過原條前端移入的細胞,在脊索的兩側形成體節中胚層。在脊索和體節中胚層上方的上胚層增厚為神經板,將來形成神經管。在原腸形成過程中,上胚層中將來形成內部器官的細胞沿原條移入,與此同時在表面的鄰近原條的細胞移向原條(圖5)。在原條充分發育之後隨著細胞不斷地進入內部,組成原條的物質越來越少,原條自前而後地逐漸變短,終至消失。在鳥類原腸形成過程中,就細胞的運動而言,原條相當於胚孔,原條的消失相當於胚孔的關閉。
 形態發生運動
形態發生運動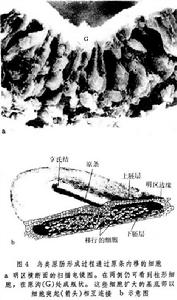 形態發生運動
形態發生運動 形態發生運動
形態發生運動器官原基形成 原腸形成之後,胚層開始發育出不同形狀的器官原基。器官原基的形成也包括形態發生運動,但往往是局限在一個胚層範圍之內,是局部的。主要是上皮細胞層的變化,它們是:
上皮細胞層的局部增厚 主要有兩種形式:①細胞增殖,如鳥類羽毛的發生,起初是在上皮層出現許多(有一定間隔)的細胞團,細胞分裂使它們繼續增長。②單個細胞的伸長。如兩棲類背部外胚層細胞在受到脊索中胚層誘導之後,由原來短柱形轉變為長柱形的細胞,而形成神經板。
細胞層的分離 細胞層可以由於在細胞間出現裂隙而分隔成層。在側板中胚層形成體避層和髒壁層時,在這二層之間,首先出現裂隙,繼而擴大為體腔。
上皮細胞層的摺疊 有各種形式:①內凹形成溝,如在神經板縱向折凹形成神經管的過程中。②外凸形成突起,某些魚類和兩棲類的外鰓是鰓區的外胚層上皮向外凸起而形成的。③內陷形成囊袋,耳囊就是主要在後腦的次級誘導下,使覆蓋其外的上皮首先增厚,再內陷形成的。
上皮細胞層的分散 上皮細胞層的分散是十分重要的形態發生運動。分散形成的間質細胞對上皮細胞的誘導刺激常常是主要的,它們可以產生次級的集聚,形成細胞團、細胞索或片狀細胞層圍繞上皮構造,以後形成結締組織和血管。很多脊椎動物的器官是由上皮和間質細胞共同形成的。譬如,來源於內胚層的上皮和來源於中胚層的間質細胞形成消化道的一些腺體,如胰腺;來源於外胚層的上皮和來源於中胚層的間質細胞發育為皮膚的衍生結構,如鱗片、羽毛等。
形態發生運動的細胞和分子基礎 不論在原腸形成過程中,或者以後的形態發生過程中,整個胚胎,或者其中的部份,發生形狀和結構變化的基本單位是細胞。單個細胞的形狀改變是細胞本身內在的變化,是形態發生運動的基礎。在原腸形成的初期,內陷開始處的細胞,形狀發生顯著變化。這些細胞的內端變圓,外端和相鄰的細胞緊密連線。並沿垂直於表面的方向伸長,結果形成具有細長勁部,球形內端的瓶狀細胞(圖6)。當這些瓶狀細胞移入內部的時候,它們把在外端和它們緊密連線的細胞一起帶入,形成表面的凹陷。這些細胞的運動決定於它們本身,因為如果把進行內陷的部分割下,離體培養,和在整體上一樣,瓶狀細胞照樣形成,而且也能移入內部,形成像胚孔一樣的凹陷。
 形態發生運動
形態發生運動在形成器官原基中,一般地說,胚胎細胞的遷移是主要的,成群的或單個的細胞脫離原來的位置,向預定的地點移動,例如神經嵴細胞的遷移,這種運動有條不紊,有時相同來源的細胞會發生分離,有時不同來源的細胞會相互聚合,按一定形式構成組織和器官。這裡既包含細胞間的識別,也包含細胞間的選擇性粘著(見細胞的親和性)。同時,遷移的途徑對於遷移中的細胞能否到達目的地,又是決定性的。這些方面了解得還很少。
