二疊紀-三疊紀滅絕事件(Permian–Triassic extinction event)是一個大規模物種滅絕事件,發生於古生代二疊紀與中生代三疊紀之間,距今大約2億5140萬年前。[1][2]若以消失的物種來計算,當時地球上70%的陸生脊椎動物,以及高達96%的海中生物消失;[3]這次滅絕事件也造成昆蟲的唯一一次大量滅絕,計有57%的科與83%的屬消失。[4][5]在滅絕事件之後,陸地與海洋的生態圈花了數百萬年才完全恢復,比其他大型滅絕事件的恢復時間更長久。[3]此次滅絕事件是地質年代的五次大型滅絕事件中,規模最龐大的一次,因此又非正式稱為大滅絕(Great Dying),或是大規模滅絕之母(Mother of all mass extinctions)。[6]
二疊紀-三疊紀滅絕事件的過程與成因仍在爭議中。[7]根據不同的研究,這次滅絕事件可分為一[1]到三[8]個階段。第一個小型高峰可能因為環境的逐漸改變,原因可能是海平面改變、海洋缺氧、盤古大陸形成引起的乾旱氣候;而後來的高峰則是迅速、劇烈的,原因可能是撞擊事件、火山爆發、或是海平面驟變,引起甲烷水合物的大量釋放。[9]
年代測定
在公元2000年之前,二疊紀與三疊紀交界的地層很少被發現,因此科學家們很難準確地估算滅絕事件的年代與經歷時間,以及影響的地理範圍。[10]在1998年,科學家研究中國浙江省長興縣煤山附近的二疊紀/三疊紀岩層,他們採用鈾-鉛測年方法,研究鋯石中的鈾/鉛比例,[2]估計二疊紀-三疊紀滅絕事件的發生年代為2億5140萬年前(誤差值為30萬年),並發現此後有隨者時間持續增高的滅絕比例。[1]在滅絕事件發生時,全球各地的碳13/碳12比例極速下降約9‰。[11][12][13][14][15]因為二疊紀/三疊紀的界線難以用放射性定年法測定,科學家們多用急遽下降的碳13/碳12比例,測定岩層中的二疊紀/三疊紀交界。[16]
在二疊紀與三疊紀交接時期,陸地與海生真菌曾有急遽的增加,原因應是當時有大量的植物與動物死亡,使得真菌得以迅速繁衍。[17]因為二疊紀/三疊紀的界線難以用放射性定年法測定,也缺乏參考用的標準化石,古動物學界一度使用真菌的繁衍高峰,測定岩層中的二疊紀/三疊紀交界。但是真菌定年法的最初提倡者,指出真菌的繁衍高峰,可能是三疊紀最早期生態系統的反覆發生現象。[17]真菌的繁衍高峰理論在最近幾年遭到許多質疑,例如:最常發現的真菌孢是Reduviasporonites,在近年被發現其實是藻類化石、 [18][11]真菌的繁衍高峰似乎不是全球性現象、[19][20]在許多地層中,真菌的繁衍高峰與二疊紀/三疊紀界線並不一致。[21]藻類被誤認為真菌胞,可能代表三疊紀時期全球各地湖泊增加,而非生物的大量死亡與腐爛。[22]
關於這次滅絕的過程、經歷時間、以及不同生物群的滅絕模式,仍然相當模糊。一些證據顯示這次滅絕事件持續約數百萬年,而在二疊紀的最後100萬年,急遽地達到高峰。[21][23]在2000年,科學家研究浙江煤山的許多富含化石的地層,藉由統計分析指出,這次滅絕事件只有一個滅亡高峰期。[1]但是,之後近年的研究指出不同的生物群在不同的時間點滅亡;舉例而言,雖然年代難以確定,介形綱(Ostracoda)與腕足動物的大規模滅絕,先後相差了72萬年到122萬年之間。[24]格陵蘭東部的一個化石保存良好的地層,動物的衰退集中在6萬到1萬年之間,而植物的衰退則持續約數十萬年之久,但也顯示滅絕事件對造成全面性的影響。[25]一個較早期的研究,認為當時有兩個滅絕高峰期,相隔500萬年,這段期間的生物滅絕高於正常標準;而最後的滅絕高峰,造成當時約80%的海洋生物滅亡,其餘海生生物則多在第一次的滅絕高峰與滅絕間期消失;近年仍有研究引用這個假設。[26]若根據此一理論,第一次的滅絕高峰發生於二疊紀瓜達鹿白階末期。[27]
瓜達鹿白階末期滅絕事件對海生生物的衝擊,隨者地區與物種的不同,而有不同程度的影響。舉例而言,腕足動物與珊瑚有嚴重的大量滅亡。[28][29]所有的恐頭獸類滅亡,僅有一屬繼續存活;[30]在有孔蟲門中,體型大型的紡錘蟲目(Fusulinida)也僅有費伯克蜓科(Verbeekinidae)繼續存活、[31]
滅絕模式
海中生物在二疊紀-三疊紀滅絕事件中,海生非脊椎動物的滅亡程度最大。在中國南部的一些富含化石的地層,在329屬的海生無脊椎動物中,有280屬在二疊紀最後兩個沉積層(以發現牙形石化石為標準)消失。[1]
根據統計分析的結果,二疊紀末海洋生物的物種數量降低,多是因為滅亡,而非物種形成的減少。[33]
在正常時期,底棲生物的滅亡率高。這次滅絕事件使底棲生物的滅亡率,達到正常時期的數倍。[34][35]以滅絕比例來看,海洋生物遭受災難性的滅亡。[36][6][1][37]
繼續存活的海生無脊椎動物則包含:腕足動物門(它們的數量在滅絕事件後持續了一段緩慢的衰退)、菊石亞綱的齒菊石目與海百合綱(幾乎滅亡,但後來再度繁盛、多樣化)。
滅亡比例最高的海洋生物,通常具有良好的循環系統、複雜的氣體交換機制、以及部分的鈣質外殼。鈣質外殼最多的物種,是滅亡比例最高的生物。[38][39]以腕足動物為例,繼續存活的物種體型較小,且是多樣化生物群落中的數量稀少成員。[40]
自從二疊紀中期(羅德階,Roadian)之後,菊石亞綱已經歷長時間的衰退,達約3000萬年之久。瓜達鹿白階(Guadalupian)末期的有限度滅絕事件,使菊石的差異性大幅減少,環境因素可能導致這次的滅絕事件。菊石的低多樣性與低差異性持續到二疊紀末滅絕事件;然而,二疊紀末滅絕事件的範圍廣泛、過程迅速。在三疊紀,菊石開始迅速多樣化,但物種之間的差異性仍然不大。[41]
在二疊紀時期,菊石的形態空間逐漸有限。在進入三疊紀的數百萬年後,菊石再度發展出原始的形態空間,但不同演化支已出現差異。[42]
陸地無脊椎動物
在二疊紀時期,陸地的昆蟲與其他無脊椎動物非常繁盛、多樣化,其中也包含地表上曾出現過的最大型昆蟲。發生在二疊紀末的滅絕事件,是目前已知惟一的昆蟲大規模滅絕事件,[4]其中有八、九個昆蟲的目消失,而至少十個目的物種減少。古網翅總目(Palaeodictyopteroidea)在二疊紀中期開始衰退,這段時間可能與植物群的變化有關係。昆蟲的衰退高峰發生於二疊紀晚期,這段時間的高峰則可能與植物群的變化(由氣候變化造成)無直接關聯。[6]
在二疊紀-三疊紀滅絕事件的前後,昆蟲的化石有極大的差異。許多古生代的昆蟲多在二疊紀-三疊紀滅絕事件後消失,只有舌鞘目、Miomoptera(內翅總目的一目)、原直翅目繼續存活到中生代。外翅總目、單尾目、古網翅總目、Protelytroptera(舌鞘目的一亞目)、原蜻蜓目在二疊紀末滅亡。在一些保存狀態良好的三疊紀晚期地層中,通常可發現與現代昆蟲族群相關的化石。[4]
陸地植物
1.植物生態系統陸地植物的化石紀錄很少見,大多數是花粉與孢子。所有大型滅絕事件對植物造成的影響,只有到科的層級。[11]若以種層級來看,減少的物種大部分可能是埋藏過程造成的假象。[11]若以優勢物種與地理分布來看,可發現植物生態系統有重大的變動。[11]
在二疊紀/三疊紀交界時,優勢的植物群改變,許多陸地植物突然開始衰退,例如荷達樹(裸子植物)與舌羊齒(種子蕨)。[43]在二疊紀/三疊紀交界之後,原本占優勢的裸子植物被石松門取代。[44]
一個針對格陵蘭東部沉積層中的花粉與孢子研究,顯示在滅絕事件發生前,當地有密集的裸子植物森林。在海生無脊椎動物群衰退時,這個大型森林也開始消失,而小型草本植物開始多樣化(包含石松門的卷柏與水韭)。裸子植物後來一度興盛,然後大量滅亡。在二疊紀末滅絕事件期間與之後,植物群發生數次興衰循環。優勢植物群從大型木本植物變動到草本植物,顯示當時許多的大型植物遭到環境改變的壓力。植物群落的接續與滅絕,發生於碳13/碳12比例極速下降之後,但兩者時間並不一致。[45]裸子植物森林的復原,花了約400萬到500萬年。[11]
2.煤層缺口在三疊紀早期的地層中,沒有發現煤礦床,而三疊紀中期的地層中,煤礦床薄,而且品質低。[46]關於煤層缺口的成因有許多假設。有科學家認為,當時有許多新的真菌、昆蟲、以及脊椎動物出現,這些具侵略性的動物造成植物的大量死亡。但是,這些動物也因二疊紀-三疊紀滅絕事件而大量滅亡,不太可能是煤層缺口的主因。[46]有可能所有可形成煤礦的植物,在這次滅絕事件中幾乎滅亡;而在1000萬年之後,新的植物才適應潮濕、酸性的泥炭沼澤,而出現新的可形成煤礦的植物。[46]非生物性因素,則包含:減少的降雨、或增加的碎屑沉積物。[11]最後,三疊紀早期的各種沉積層很少被發現,煤層的缺口可能反映了沉積層的稀少狀況。有可能形成煤礦的三疊紀早期生態系統,目前還沒有被發現。[11]例如在澳大利亞東部,當時可能長時間處在寒冷的氣候,這種天氣狀況適合泥炭的形成。在可以形成泥炭的植物中,有95%在這次滅絕事件中消失。[47]但是,澳大利亞與南極洲的煤礦床在P-Tr事件發生前,就已經消失。[11]
3.陸地脊椎動物在二疊紀末期,許多陸地脊椎動物消失,而某些演化支甚至幾乎滅亡。某些存活的族系,並未長時間繼續存在,而其他族系則開始多樣化,並長期存活者。有充足證據顯示,超過2/3的陸生兩棲動物、蜥形綱、獸孔目的科在這個滅絕事件中消失。大型的草食性動物遭受嚴重的打擊。除了前棱蜥科,幾乎所有的二疊紀無孔亞綱動物滅亡;龜鱉目具有無孔類的頭骨,被認為可能是從前棱蜥科的近親演化而來。盤龍目早在二疊紀末期前就已滅亡。由於二疊紀的雙孔亞綱化石很少,無法確定滅絕事件對雙孔亞綱所造成的影響;雙孔亞綱包含大部分的爬行動物,例如蜥蜴、蛇、鱷魚、恐龍、以及恐龍的後代鳥類。[48][49]
滅絕模式的可能解釋
滅亡程度最嚴重的海生動物,包含:可以製造鈣質外殼(例如碳酸鈣)、代謝率低、以及呼吸系統較弱的物種,尤其是鈣質海綿類、四射珊瑚、床板珊瑚、具鈣質外殼的腕足動物、苔蘚動物門、棘皮動物門,上述動物中約有81%的屬滅亡。而無法製造鈣質外殼的近親,受到的影響較小,例如海葵(演化出現代的珊瑚)。具有高代謝率、呼吸系統良好、以及缺乏鈣質外殼的動物,滅亡的幅度較小;牙形石是個例外,約有33%的屬消失。[50]這種滅絕模式,與組織缺氧產生的效應相符合。在當時,幾乎所有的大陸棚海域都有嚴重的缺氧現象,但缺氧現象無法解釋有限度的滅絕模式;因此缺氧不是造成海生生物滅絕的唯一成因。一個針對二疊紀晚期到三疊紀早期大氣層的數學模型,指出在這段期間,大氣層中的氧氣有明顯、長時期的衰退;這個衰退不因二疊紀/三疊紀交界而增加,並在三疊紀早期達到最低點,但仍高於現今的比例。由此得知,氧氣比例的衰退與滅絕模式並不一致。[50]
另一方面,海生生物的滅絕模式也與高碳酸血症(二氧化碳過高)產生的效應相符合。當二氧化碳濃度高於正常時,將對生物造成傷害:降低呼吸色素攜帶氧到組織的能力、使體液更為酸性、妨礙動物形成碳酸鈣外殼;當濃度更高時,會使生物進入昏迷狀態。除了上述反應,二氧化碳濃度高,會使生物製造鈣質外殼更加困難。與陸地生物相比,海洋生物對於二氧化碳濃度的變化相當敏感。溶解於水中的二氧化碳,是氧的28倍。對於進行呼吸作用的動物,二氧化碳的排除,與氣體經過呼吸系統膜相關;所以海洋動物會將身體內的二氧化碳保持在低濃度,低於陸地動物。若二氧化碳濃度增加,會妨礙海洋動物合成蛋白質、降低受精機率、形成不完整的鈣質外殼。[50]
目前很少發現跨越二疊紀與三疊紀交界的陸相地層,因此難以詳細研究陸地生物的滅絕/存活比例。二疊紀與三疊紀的昆蟲有顯著的不同,但是在二疊紀晚期到三疊紀早期,有個約1500萬年的昆蟲化石斷層。目前最足以研究二疊紀/三疊紀陸地脊椎動物變化的地層,位於南非的卡魯盆地(karoo);但目前仍沒有足夠的研究可供結論。[50]
生態系統的復原
早期的研究認為在二疊紀末滅絕事件後,地表的生物迅速的復原,但這些研究多是針對在滅絕存活的先驅生物(Pioneer organisms)的研究結果,例如水龍獸。近年的研究認為,當時經歷相當長的時間,生態系統才恢復以往的高度多樣性、複雜的食物鏈、與眾多生態位。在滅絕事件後,發生數次的滅絕波動,限制棲息地的復原,因此生態系統花了長時期才完全復原。近年的研究指出,地表生態系統直到進入三疊紀中期,約是滅絕事件後400萬到600萬年,才開始復原。[51]某些研究人員更指出,直到滅絕事件後3000萬年,進入三疊紀晚期,地表生態系統才完全復原。[52]
 懷馬卡里里河,分布於紐西蘭南島,屬於辮狀河
懷馬卡里里河,分布於紐西蘭南島,屬於辮狀河懷馬卡里里河,分布於紐西蘭南島,屬於辮狀河在三疊紀早期,約是滅絕事件後400萬到600萬年,陸地的植物數量不足,造成煤層缺口,使當時的草食性動物面臨食物短缺。[46]南非卡魯盆地的河流,從曲折河改變為辮狀河,顯示該地區的植被曾長時間非常稀少。[53]
三疊紀早期的主要生態系統,無論是植物、動物,或是海洋、陸地系統,優勢生物都是少數特定、且全球分布的物種。舉例而言,獸孔目的水龍獸,這種草食性動物的數量,占了當時陸地草食性動物約90%;以及雙殼綱的克氏蛤(Claraia)、正海扇(Eumorphotis)、蚌形蛤(Unionites )、Promylina。正常的生態系統應由眾多物種構成,並占據者不同的棲息地與生態位。[43][54]
在滅絕事件存活下來的某些劣勢物種,數量與棲息地大幅增加。例如:雙殼綱的海豆芽(Lingula)、出現於奧陶紀的疊層石、石松門的肋木(Pleuromeia)、種子蕨的二叉羊齒(Dicrodium)。[9][55][54]
海洋生態系統的改變
在二疊紀末滅絕事件後,濾食性固著動物(例如海百合)的數量大幅減少在二疊紀-三疊紀滅絕事件發生前,接近67%
 在二疊紀末滅絕事件後,濾食性固著動物(例如海百合)的數量大幅減少
在二疊紀末滅絕事件後,濾食性固著動物(例如海百合)的數量大幅減少的海生動物是固著動物,附著于海床生存;在中生代期間,只有50%的海生動物是固著動物,其餘皆是可自由行動的動物。研究顯示,事件發生後,棲息在海底表層的濾食性固著動物,例如腕足動物、海百合,物種數量減少;而構造較複雜的動物,例如蝸牛、海膽、螃蟹,物種數量增加。
在二疊紀末滅絕事件發生前,同時存在複雜與單純的海洋生態系統,比例相當;生態系統經過復原後,複雜的海洋生物群落在數量上超過單純的海洋生物群落,比例相當於3/1;[56]這使海生掠食者面臨食物競爭的壓力,促成中生代的海洋生物進一步演化。
在滅絕事件發生前,雙殼綱是非常罕見的物種。在三疊紀,雙殼綱成為數量眾多、多樣化的生物群;其中的厚殼蛤(Rudists),是形成中生代時期礁岩的主要生物。部分研究人員認為雙殼綱的興起,發生於兩次滅絕高峰的間期。[57]
在滅絕事件中,海百合綱的物種數量、多樣性大幅減少,但沒有完全滅亡。[58]在滅絕事件後,海百合綱接者發生適應輻射,除了數量、棲息地增加,也發展出柔軟的腕足、具可動性。[59]
陸地脊椎動物
在三疊紀早期,水龍獸是陸地上最繁盛的脊椎動物水龍獸屬於獸孔目二齒獸類,是種體型接近豬的草食性動物,在三
 在三疊紀早期,水龍獸是陸地上最繁盛的脊椎動物
在三疊紀早期,水龍獸是陸地上最繁盛的脊椎動物疊紀最早幾個時期,占據了90%的陸地動物生態位。[9]除此之外,還有體型較小的肉食性犬齒獸類倖存,犬齒獸類是哺乳動物的祖先。
在三疊紀的早期,主龍類的數量遠少於獸孔目,但它們在三疊紀中期,取代獸孔目的生態位。[9]在三疊紀中到晚期,主龍類的恐龍演化出現,並逐漸成為中生代的優勢陸地動物。[60]獸孔目與主龍類的興衰,與哺乳類的演化過程有關。獸孔目與隨後出現的哺乳形類,在這個時期多半演化成小型、夜行性的食蟲動物。夜間的生活習性,至少使哺乳形類發展出毛髮與較高的代謝率。[61]
兩棲動物離片椎目的某些物種,在滅絕事件中幾乎滅亡,但快速復原。在三疊紀的大部分時間,蝦蟆螈(Mastodonsaurus)與長吻迷齒螈下目(Trematosauria)是主要的水生與半水生掠食動物,以其他四足類、魚類為食。[62]
陸地脊椎動物經過相當長的時間,才從二疊紀末滅絕事件復原。一位研究人員估計陸地脊椎動物花了3000萬年,直到三疊紀晚期,才恢復之前的繁盛與多樣性。此時的陸地脊椎動物包含:兩棲動物、原始主龍類、恐龍、翼龍目、早期鱷魚、以及哺乳形類。[3]
滅絕原因
關於二疊紀-三疊紀滅絕事件的發生過程,目前已有多種假設,包含劇烈與緩慢的過程;白堊紀-第三紀滅絕事件的發生過程,也有類似的假設。劇烈過程理論的成因,包含大型或多顆隕石造成的撞擊事件、火山爆發、或是海床急驟釋放出大量甲烷水合物。緩慢過程理論的成因,包含海平面改變、缺氧、以及逐漸增加的乾旱氣候。[9]
1.撞擊事件
位於南極洲的威爾克斯地隕石坑,直徑達500公里,形成時間在5億年內,規模與時代使它成為本次滅絕事件的可疑成因,但證據不夠明確。白堊紀-第三紀滅絕事件由撞擊事件造成的證據,促使科學家們推論其他滅絕事件由撞擊事件造的的可能性,尤其是二疊紀-三疊紀滅絕事件。因此,科學家們盡力尋找那個時代的大型隕石坑與撞擊證據。
目前已在部分二疊紀/三疊紀交界的地層,發現撞擊事件的證據,例如:在澳大利亞與南極洲發現罕見的衝擊石英、[63][64]富勒烯包覆的外太空惰性氣體、[65]南極洲發現的玻璃隕石、[66]地層中常見鐵、鎳、矽微粒。 [67]但是,上述證據的真實性多受到懷疑。[68][69][70][71]在南極洲石墨峰(Graphite Peak)發現的衝擊石英,經過光學顯微鏡與穿透式電子顯微鏡重新檢驗後,發現其中的結構並非由撞擊產生,而是形變與地殼活動(例如火山)造成的。[72]
目前已經發現數個可能與二疊紀末滅絕事件有關的隕石坑,包含:澳大利亞西北外海的貝德奧高地、[64]南極洲東部的威爾克斯地隕石坑。[73][74]但沒有可信服的證據,可證明這兩個地形是由撞擊產生。以威爾克斯地隕石坑為例,這個位在冰原下的凹地,年代無法確定,可能晚於二疊紀末滅絕事件才形成。
如果二疊紀末滅絕事件的主因是由於撞擊,很有可能隕石坑已經消失於地表。地球表面有70%是海洋,所以隕石或彗星撞擊海洋的機率,是撞擊陸地的兩倍以上。但是,地球的海洋地殼會因聚合與隱沒作用而消失於地表,所以目前無法找到距今2億年以上的海洋地殼。如果當時有非常大型的撞擊事件,撞擊會使該處地殼破裂、變薄,造成大量的熔岩。[75]
撞擊理論最受到關注的原因是,它可與其他滅絕現象產生因果連結,[76]例如西伯利亞地盾火山爆發,可能由大型撞擊產生,[77]甚至是大型撞擊的對蹠點。[78][76]即使撞擊發生於海洋,隕石坑因為隱沒帶而消失於地表,應會留下其他證據。如同白堊紀-第三紀滅絕事件,如果有撞擊事件發生,會產生大量撞擊拋出物,相同時代的許多地層會發現大量親鐵元素,例如銥。若滅絕事件由隕石撞擊引發,可以解釋在滅絕後,生物沒有快速的適應演化。
2.火山爆發
接近二疊紀/三疊紀交界時的全球地圖。西伯利亞地盾火山爆發位於北方淺海的東岸。峨嵋山地盾火山爆發時間較
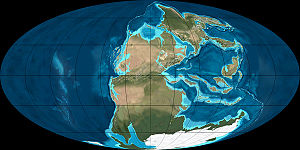 接近二疊紀/三疊紀交界時的全球地圖。西伯利亞地盾火山爆發位於北方淺海的東岸。峨嵋山地盾火山爆發時間較早,位置在東方大型獨立島嶼。
接近二疊紀/三疊紀交界時的全球地圖。西伯利亞地盾火山爆發位於北方淺海的東岸。峨嵋山地盾火山爆發時間較早,位置在東方大型獨立島嶼。早,位置在東方大型獨立島嶼。
西伯利亞地盾的熔岩範圍在二疊紀的最後一期,發生兩個大規模火山爆發:西伯利亞地盾、峨嵋山地盾。峨嵋山地盾位於現今中國四川省,規模較小,形成時間是瓜達鹿白階末期,形成時的位置接近赤道。[79]西伯利亞地盾火山爆發是地質史上已知最大型的火山爆發之一,熔岩面積達200萬平方公里。西伯利亞地盾火山爆發原本被認為持續約數百萬年,但近年的研究認為西伯利亞地盾形成於2億5120萬年前(誤差值為30萬年),在二疊紀末期。[1][80]
峨嵋山地盾與西伯利亞地盾火山爆發,可能製造大量灰塵與酸性微粒,遮蔽照射到地表的陽光,妨礙陸地與海洋透光帶的生物進行光合作用,進而遭成食物鏈的崩潰。大氣層中的酸性微粒,最後形成酸雨降落到地表。酸雨對陸地植物、可製造碳酸鈣硬殼的軟體動物與浮游生物造成傷害。火山爆發也釋放大量二氧化碳,形成全球暖化。大氣層中的灰塵與酸性物質降落到地表之後,過量的二氧化碳持續形成溫室效應。[76]
與其他火山相比,西伯利亞地盾更為危險。洪流玄武岩會產生大量的流動性熔岩,只會噴發少量的蒸氣、碎屑進入大氣層。但是,西伯利亞地盾火山爆發的噴出物質,似乎有20%是火山碎屑,這些火山灰與火山礫進入大氣層後,會造成短時期的氣候寒冷。[81]帶有玄武岩的熔岩侵入碳酸鹽岩、或帶有大型煤層的地區,會產生大量的二氧化碳,會在大氣層的灰塵降落到地表後,造成全球暖化。[76]
這些火山爆發事件的規模是否足以造成二疊紀末滅絕事件,仍有爭議。峨嵋山地盾接近赤道區,火山爆發所製造的灰塵與酸性物質,會對全世界造成影響。西伯利亞地盾的規模較大,但位置在北極區內,或在北極區附近。如果西伯利亞地盾火山爆發的持續時間在20萬年內,會使大氣層中的二氧化碳達到正常程度的兩倍。近年的氣候模型顯示,大氣層中的二氧化碳含量加倍,會使全球氣候上升1.5°C到4.5°C,這會造成嚴重的影響,但沒有到二疊紀-三疊紀滅絕事件的嚴重程度。[76]
在2005年,日本NHK與加拿大國家電影委員會(NFB)製作的《地球大紀行》(Miracle Planet)電視節目,提出火山爆發形成的輕微全球暖化,導致甲烷水合物的氣化;由於甲烷氣體對全球暖化的影響,是二氧化碳的45倍,甲烷水合物的氣化進而導致不斷循環的全球暖化。
3.甲烷水合物的氣化
科學家們已在全球許多地點的二疊紀末碳酸鹽礦層中,發現當時的碳13/碳12比例有迅速減少的跡象,減少了約10‰左右[82][37]從二疊紀末開始,碳13/碳12比例發生了一系列的上升與下降現象,直到三疊紀中期才穩定、停止;而發生於二疊紀末的第一次變動,是其中規模最大、最迅速的變動。在三疊紀中期,碳13/碳12比例穩定之後,可製造碳酸鈣的動物開始復原。[38]
碳13/碳12比例的下降,可能有以下多種因素:[83]
 甲烷水合物即俗稱的可燃冰,大部分位在大陸棚,被視為未來的潛在能源之一。
甲烷水合物即俗稱的可燃冰,大部分位在大陸棚,被視為未來的潛在能源之一。火山爆發產生的氣體,其碳13/碳12比例低於正常值約5~8‰。但若要使全球的碳13/碳12比例下降約10‰,其火山爆發的規模將超越目前已知的任何地質紀錄。[84]
生物活動降低,使環境中的碳12更慢被攝取,更多碳12進入沉積層,而使碳13/碳12比例降低。所有的化學反應是建立在原子間的電磁力。較輕的同位素,其化學反應較快。所以生物化學過程會用到較輕的同位素。但關於古新世-始新世交替時期最大熱量(PETM)事件的研究發現,該時期的碳13/碳12比例小幅降低約3到4‰。即使將全部的有機碳(包含生物、土壤、海洋)進入土壤沉積層中,也不會達到古新世/始新世交替時的小幅度碳13/碳12比例下降。由此可知生物活動降低不是二疊紀末期的碳13/碳12比例降低的原因。[84]
埋在沉積層中的死亡生物,體內的碳13/碳12比例小於正常值約20到25‰。就理論上而言,如果海平面迅速降低,淺海地區的沉積層曝露到空氣後,開始氧化作用。但若要使全球的碳13/碳12比例下降約10‰,要有6.5到8.4兆噸有機碳經氧化後形成,而沉積層本身需要數十萬年的氧化。這似乎不太可能發生。[6]
間歇性海洋高氧與缺氧事件,也可能是三疊紀早期的碳13/碳12比例下降的原因。[38]全球性的海洋缺氧現象,本身也是滅絕事件的原因之一。二疊紀末期到三疊紀早期的陸地,多為熱帶地區。熱帶的大型河流會將沉積層中有有機碳帶入海洋,尤其是低緯度的鄰近海盆。生物化學過程會用到較輕的同位素,所以有機碳的碳13/碳12比例低。大量有機碳的迅速(以相對而言)釋放與沉降,可能會引發間歇性的高氧/缺氧事件。[85]元古宙晚期到寒武紀交接時期,也曾發生碳13/碳12比例的下降,可能與此相關,或是其他與海洋相關的因素。[38]
甲烷水合物即俗稱的可燃冰,大部分位在大陸棚,被視為未來的潛在能源之一。其他的理論則有:海洋大量釋放二氧化碳、[86]以及全球的碳循環系統經歷長時間的重整。[83]
但最有可能導致全球性碳13/碳12比例下降的因素,是甲烷水合物氣化產生的甲烷。[6]而利用碳循環模型模擬的結果,甲烷最有可能導致如此大幅的下降。[83][86]甲烷水合物是固態形式的水於晶格(水合物)中包含大量的甲烷。甲烷是由甲烷菌製造,碳13/碳12比例低於正常值約60‰。甲烷在特定的壓力與溫度下,會形成包合物,例如永凍層的近表層,並在大陸棚、更深的海床等地區大量形成。甲烷水合物通常出現在海平面300米以下的沉積層。最深可在水深2000米處發現,淡大部分在水深1100米以上。[87]
西伯利亞地盾火山爆發產生的熔岩面積,是以往認定的兩倍以上,新發現的熔岩地區,在二疊紀末時幾乎是淺海。極有可能這些淺海地區蘊含甲烷水合物,熔岩流入海床後,促使甲烷水合物的汽化。[88]
由於甲烷本身是種非常強的溫室氣體,大量的甲烷被視為造成全球暖化的主要原因。證據顯示該時期的全球氣溫上升,赤道區上升約6°C,高緯度地區上升更多。例如:氧18/氧16比例的下降、[89]舌羊齒植物群(舌羊齒與生存於相同地區的植物)消失,由生存於低緯度的植物群取代。[90][9]
碳13/碳12比例的變動,被認為與大量釋放的甲烷有關,但兩者在三疊紀早期的變動模式,並不吻合。要造成如此大的氣候變遷,所需要的甲烷量,是引發古新世-始新世交替時期最大熱量的五倍。[38]但三疊紀早期曾出現大量碳13,使碳13/碳12比例迅速增加,而後下降。[38]
4.海平面改變
當原本沉浸的海床露出海平面時,會造成海退。海平面的下降會使淺海的生存區域減少,破壞當地的生態系統。淺海的可棲息地,富含食物鏈下層的生物,這些生物的減少,使賴其以維生的生物競爭食物更激烈。[91]海退與滅絕事件之間似乎有部份關聯,但另有證據認為兩者間沒有關係,而海退會形成新的棲息地。[9]海平面的變化,同時也導致海底沉積物的變化,並影響海水溫度與鹽度,進一步造成海生生物的多樣性衰退。[92]
5.海洋缺氧
有證據顯示,二疊紀末期的海洋發生了缺氧事件。在格陵蘭東部的一個二疊紀末期海相沉積層,指出當時有明顯、快速的海洋缺氧現象。[93]而數個二疊紀末沉積層的鈾/釷比例,也指出在這次滅絕事件發生時,海洋有嚴重的缺氧現象。[94]
缺氧事件可能導致海洋生物的大量死亡,只有棲息于海底泥層、可以進行缺氧呼吸的細菌不受影響。另有證據顯示,這次海洋缺氧事件,造成海床大量釋放硫化氫。
海洋缺氧事件的原因,可能是長時間的全球暖化,降低赤道區與極區之間的溫度梯度,進而造成溫鹽環流系統的緩慢,甚至停止。溫鹽環流系統的緩慢或停止,可能使得海洋中的含氧量減少。[94]
但是,某些研究人員架構出二疊紀末期的海洋溫鹽環流系統,認為當時的溫鹽環流系統無法解釋深海區域的缺氧現象。[95]
6.硫化氫
二疊紀末期發生的海洋缺氧事件,可能使硫酸鹽還原菌成為海洋生態系統中的優勢物種,包含脫硫桿菌目、脫硫弧菌目、互營桿菌目、熱脫硫桿菌門,這些生物會製造大量的硫化氫,過量的硫化氫會對陸地、海洋中的動植物造成毒害,並破壞臭氧層,使生物暴露在紫外線下。[96]在二疊紀末到三疊紀早期發現許多綠菌,它們進行不產氧光合作用,釋放出硫化氫。綠菌的興盛時期,與二疊紀末滅絕事件和事後的長期復原,時期相符。[97]大氣層中的二氧化碳增加,植物卻大規模滅亡,硫化氫理論可以解釋植物的大規模滅亡。二疊紀末地層中的孢子化石,多數帶有不正常特徵,可能是由硫化氫破壞臭氧層,大量的紫外線進入地表造成。
盤古大陸的地圖
盤古大陸的形成
在二疊紀中期(約是烏拉爾世的空谷階),幾乎地表的所有大陸聚合成盤古大陸,盤古大陸由泛大洋環繞者,而東亞部份直到二疊紀末期才與盤古大陸聚合。[98]盤古大陸的形成,使得全球大部分的淺水區域消失,而淺水區域是海洋中最多生物棲息的部份。原本隔離的大陸架連線之後,使彼此獨立的生態系統開始互相競爭。盤古大陸的形成,造成了單一的海洋循環系統,以及單一的大氣氣候系統,在盤古大陸的海岸形成季風氣候,而廣大的內陸則形成乾旱的氣候。
在盤古大陸形成後,海洋生物數量減少,滅亡比例接近其他大型滅絕事件。盤古大陸的形成,似乎對陸地生物沒有造成嚴重的變化,二疊紀晚期的獸孔目反而因此擴大生存領域、更為多樣性。盤古大陸的形成,使海洋生物開始減少,但不是導致二疊紀末滅絕事件的直接原因。
多重原因
二疊紀-三疊紀末滅絕事件的產生原因,可能由上述事件連鎖、交錯形成,並日趨嚴重。西伯利亞地盾的火山爆發,除了產生大量的二氧化碳與甲烷,也破壞鄰近地區的煤層與大陸架。接下來的全球暖化,間接導致地質歷史中最嚴重的海洋缺氧事件。海洋的缺氧,使綠菌等進行進行不產氧光合作用生物的興起,它們釋放出大量的硫化氫。
但是,這連鎖、交錯的事件,部分環節相當薄弱。碳13/碳12比例的變動,被認為與大量釋放的甲烷有關,但兩者在三疊紀早期的變動模式,並不吻合。[38]二疊紀末期的海洋溫鹽環流系統,不會造成深海區域的缺氧事件。[95]
注釋
1.^ 1.0 1.1 1.2 1.3 1.4 1.5 1.6 (英文) Jin YG, Wang Y, Wang W, Shang QH, Cao CQ, Erwin DH(2009年5月8日).Pattern of Marine Mass Extinction Near the Permian–Triassic Boundary in South China.Science,289(5478):432–436.doi:10.1126/science.289.5478.432.PMID 10903200.
2.^ 2.0 2.1 (英文)Bowring SA, Erwin DH, Jin YG, Martin MW, Davidek K, Wang W(1998年5月8日).U/Pb Zircon Geochronology and Tempo of the End-Permian Mass Extinction.Science,280(1039):1039–1045.doi:10.1126/science.280.5366.1039.
3.^ 3.0 3.1 3.2 (英文)Benton M J(2005).When Life Nearly Died: The Greatest Mass Extinction of All Time.Thames & Hudson.ISBN 978-0500285732.
4.^ 4.0 4.1 4.2 (英文)Labandeira CC, Sepkoski JJ(1993年).Insect diversity in the fossil record.Science,261(5119):310–5.PMID 11536548.於2008年1月8日查閱.
5.^ (英文)Sole, R. V., and Newman, M., 2002. "Extinctions and Biodiversity in the Fossil Record - Volume Two, The earth system: biological and ecological dimensions of global environment change" pp. 297-391, Encyclopedia of Global Enviromental Change John Wilely & Sons.
6.^ 6.0 6.1 6.2 6.3 6.4 (英文)Erwin DH(1993).The great Paleozoic crisis; Life and death in the Permian.Columbia University Press.ISBN 0231074670.
7.^ (英文)Yin H, Zhang K, Tong J, Yang Z, Wu S.The Global Stratotype Section and Point (GSSP) of the Permian-Triassic Boundary.Episodes,24(2):102–114.
8.^ (英文)Yin HF, Sweets WC, Yang ZY, Dickins JM,.Permo-Triassic Events in the Eastern Tethys.Cambridge Univ. Pres, Cambridge, 1992.
9.^ 9.0 9.1 9.2 9.3 9.4 9.5 9.6 (英文)Tanner LH, Lucas SG & Chapman MG(2004年).Assessing the record and causes of Late Triassic extinctions(PDF).Earth-Science Reviews,65(1-2):103-139.doi:10.1016/S0012-8252(03)00082-5.於2007年10月22日查閱.
10.^ (英文)Erwin, D.H(1993).The Great Paleozoic Crisis: Life and Death in the Permian.New York isbn=0231074670:Columbia University Press.
11.^ 11.0 11.1 11.2 11.3 11.4 11.5 11.6 11.7 11.8 (英文)McElwain, J.C.; Punyasena, S.W.(2007年).Mass extinction events and the plant fossil record.Trends in Ecology & Evolution,22(10):548–557.doi:10.1016/j.tree.2007.09.003.
12.^ (英文)Magaritz, M(1989年5月8日).13C minima follow extinction events: a clue to faunal radiation.Geology,17:337–340.
13.^ (英文)Krull, S.J., and Retallack, J.R.(2009年5月8日).13C depth profiles from paleosols across the Permian–Triassic boundary: Evidence for methane release.GSA Bulletin,112(9):1459–1472.doi:[[doi:10.1130%2F0016-7606%282000%29112%253C1459%3ACDPFPA%253E2.0.CO%3B2|10.1130/0016-7606(2000)112%3C1459:CDPFPA%3E2.0.CO;2]].
14.^ (英文)Dolenec, T., Lojen, S., Ramovs, A.(2009年5月8日).The Permian–Triassic boundary in Western Slovenia (Idrijca Valley section): magnetostratigraphy, stable isotopes, and elemental variations.Chemical Geology,175(1):175–190.doi:10.1016/S0009-2541(00)00368-5.
15.^ (英文)Musashi, M., Isozaki, Y., Koike, T. and Kreulen, R.(2009年5月8日).Stable carbon isotope signature in mid-Panthalassa shallow-water carbonates across the Permo–Triassic boundary: evidence for 13C-depleted ocean.Earth Planet. Sci. Lett.,193:9–20.doi:10.1016/S0012-821X(01)00398-3.
16.^ (英文)Dolenec, T., Lojen, S., and Ramovs, A.(2009年5月8日).The Permian-Triassic boundary in Western Slovenia (Idrijca Valley section): magnetostratigraphy, stable isotopes, and elemental variations.Chemical Geology,175:175–190.doi:10.1016/S0009-2541(00)00368-5.
17.^ 17.0 17.1 (英文)H Visscher, H Brinkhuis, D L Dilcher, W C Elsik, Y Eshet, C V Looy, M R Rampino, and A Traverse(1996年5月8日).The terminal Paleozoic fungal event: Evidence of terrestrial ecosystem destabilization and collapse.Proceedings of the National Academy of Sciences,93(5):2155–2158.於2007年9月20日查閱.
18.^ (英文)Foster, C.B.; Stephenson, M.H.; Marshall, C.; Logan, G.A.; Greenwood, P.F.(2002年).A Revision Of Reduviasporonites Wilson 1962: Description, Illustration, Comparison And Biological Affinities.Palynology,26(1):35–58.doi:10.2113/0260035.
19.^ (英文)López-Gómez, J. and Taylor, E.L.(2009年5月8日).Permian-Triassic Transition in Spain: A multidisciplinary approach.Palaeogeography, Palaeoclimatology, Palaeoecology,229(1-2):1–2.doi:10.1016/j.palaeo.2005.06.028.
20.^ (英文)Looy, C.V.; Twitchett, R.J.; Dilcher, D.L.; Van Konijnenburg-van Cittert, J.H.A.; Visscher, H.(2005年).Life in the end-Permian dead zone.Proceedings of the National Academy of Sciences,162(4):653–659.doi:10.1073/pnas.131218098.PMID 11427710.“See image 2”
21.^ 21.0 21.1 (英文)Ward PD, Botha J, Buick R, De Kock MO, Erwin DH, garrison GH, Kirschvink JL & Smith R(2009年5月8日).Abrupt and Gradual Extinction Among Late Permian Land vertebrates in the Karoo Basin, South Africa.Science,307(5710):709–714.doi:10.1126/science.1107068.
22.^ (英文)Retallack, G.J.; Smith, R.M.H.; Ward, P.D.(2003年).Vertebrate extinction across Permian-Triassic boundary in Karoo Basin, South Africa.Bulletin of the Geological Society of America,115(9):1133–1152.doi:10.1130/B25215.1.
23.^ (英文)Rampino MR, Prokoph A & Adler A(2009年5月8日).Tempo of the end-Permian event: High-resolution cyclostratigraphy at the Permian–Triassic boundary.Geology,28(7):643–646.doi:[[doi:10.1130%252F0091-7613%25282000%252928%253C643%253ATOTEEH%253E2.0.CO%253B2|10.1130%2F0091-7613%282000%2928%3C643%3ATOTEEH%3E2.0.CO%3B2]].
24.^ (英文)Wang, S.C.; Everson, P.J.(2007年).Confidence intervals for pulsed mass extinction events.Paleobiology,33(2):324–336.doi:10.1666/06056.1.
25.^ (英文)Twitchett RJ Looy CV Morante R Visscher H & Wignall PB(2009年5月8日).Rapid and synchronous collapse of marine and terrestrial ecosystems during the end-Permian biotic crisis.Geology,29(4):351–354.doi:[[doi:10.1130%2F0091-7613%282001%29029%3C0351%3ARASCOM%3E2.0.CO%3B2|10.1130/0091-7613(2001)029<0351:RASCOM>2.0.CO;2]].
26.^ Retallack, G.J.; Metzger, C.A.; Greaver, T.; Jahren, A.H.; Smith, R.M.H.; Sheldon, n.d.(2006年).Middle-Late Permian mass extinction on land.Bulletin of the Geological Society of America,118(11-12):1398–1411.doi:10.1130/B26011.1.
27.^ (英文)Stanley SM & Yang X(1994年5月8日).A Double Mass Extinction at the End of the Paleozoic Era.Science,266(5189):1340–1344.doi:10.1126/science.266.5189.1340.PMID 17772839.
28.^ (英文)Shen, S., and Shi, G.R.(2009年5月8日).Paleobiogeographical extinction patterns of Permian brachiopods in the Asian-western Pacific region.Paleobiology,28:449–463.doi:[[doi:10.1666%2F0094-8373%282002%29028%3C0449%3APEPOPB%3E2.0.CO%3B2|10.1666/0094-8373(2002)028<0449:PEPOPB>2.0.CO;2]].
29.^ (英文)Wang, X-D, and Sugiyama, T.(2000年12月1日).Diversity and extinction patterns of Permian coral faunas of China.Lethaia,33(4):285–294.doi:10.1080/002411600750053853.
30.^ Retallack, G.J., Metzger, C.A., Jahren, A.H., Greaver, T., Smith, R.M.H., and Sheldon, N.D(November/December 2006).Middle-Late Permian mass extinction on land.GSA Bulletin,118(11/12):1398–1411.doi:10.1130/B26011.1.
31.^ (英文)Ota, A, and Isozaki, Y.(2006年3月1日).Fusuline biotic turnover across the Guadalupian–Lopingian (Middle–Upper Permian) boundary in mid-oceanic carbonate buildups: Biostratigraphy of accreted limestone in Japan.Journal of Asian Earth Sciences,26(3-4):353-368.
32.^ (英文) Racki G(1999年5月8日).Silica-secreting biota and mass extinctions: survival processes and patterns,154:107–132.doi:10.1016/S0031-0182(99)00089-9.
33.^ Bambach, R.K.; A.H. Knoll & S.C. Wang (December 2004), "Origination, extinction, and mass depletions of marine diversity", Paleobiology 30 (4): 522–542
34.^ (英文)Stanley, S.M.(2008年).Predation defeats competition on the seafloor.Paleobiology,34(1):1–21.doi:10.1666/07026.1.於2008年5月13日查閱.
35.^ (英文)Stanley, S.M.(2007年).An Analysis of the History of Marine Animal Diversity.Paleobiology,33(sp6):1–55.doi:10.1666/06020.1.
36.^ (英文)McKinney, M.L.(1987年5月8日).taxonomic selectivity and continuous variation in mass and background extinctions of marine taxa.Nature,325(6100):143–145.doi:10.1038/325143a0.
37.^ 37.0 37.1 (英文)Twitchett RJ, Looy CV, Morante R, Visscher H, Wignall PB(2009年5月8日).Rapid and synchronous collapse of marine and terrestrial ecosystems during the end-Permian biotic crisis.Geology,29(4):351–354.doi:[[doi:10.1130%2F0091-7613%282001%29029%253C0351%3ARASCOM%253E2.0.CO%3B2|10.1130/0091-7613(2001)029%3C0351:RASCOM%3E2.0.CO;2]].
38.^ 38.0 38.1 38.2 38.3 38.4 38.5 38.6 (英文)Payne, J.L.; Lehrmann, D.J.; Wei, J.; Orchard, M.J.; Schrag, D.P.; Knoll, A.H.(2004年).Large Perturbations of the Carbon Cycle During Recovery from the End-Permian Extinction.Science,305(5683):506.doi:10.1126/science.1097023.PMID 15273391.
39.^ (英文)Knoll, A.H.; Bambach, R.K.; Canfield, D.E.; Grotzinger, J.P.(1996年).Comparative Earth history and Late Permian mass extinction.Science(Washington),273(5274):452–456.doi:10.1126/science.273.5274.452.PMID 8662528.
40.^ (英文)Leighton, L.R.; Schneider, C.L.(2008年).Taxon characteristics that promote survivorship through the Permian–Triassic interval: transition from the Paleozoic to the Mesozoic brachiopod fauna.Paleobiology,34(1):65–79.doi:10.1666/06082.1.
42.^ (英文)Villier, L.(2004年).Morphological disparity of Ammonoids and the Mark of Permian Mass Extinctions.Science,306:264.doi:10.1126/science.1102127.
42.^ (英文)Saunders, W. B.(2008年).Morphologic and taxonomic history of Paleozoic ammonoids in time and morphospace.Paleobiology,34:128.doi:10.1666/07053.1.
43.^ 43.0 43.1 (英文)Retallack, GJ(1995年5月8日).Permian–Triassic life crisis on land.Science,267(5194):77–80.doi:10.1126/science.267.5194.77.
44.^ (英文)Looy, CV Brugman WA Dilcher DL & Visscher H(1999年5月8日).The delayed resurgence of equatorial forests after the Permian–Triassic ecologic crisis.Proceedings National Academy of Sciences,96:13857–13862.PMID 10570163.
45.^ (英文)Looy, CV; Twitchett RJ, Dilcher DL, &Van Konijnenburg-Van Cittert JHA and Henk Visscher.(2001年7月3日).Life in the end-Permian dead zone.Proceedings of the National Academy of Sciences,14(98):7879–7883.doi:10.1073/pnas.131218098.
46.^ 46.0 46.1 46.2 46.3 (英文)Retallack GJ Veevers JJ & Morante R(1996年5月8日).Global coal gap between Permian–Triassic extinctions and middle Triassic recovery of peat forming plants.GSA Bulletin,108(2):195–207.於2007年9月29日查閱.
47.^ (英文)Michaelsen P(2009年5月8日).Mass extinction of peat-forming plants and the effect on fluvial styles across the Permian–Triassic boundary, northern Bowen Basin, Australia.Palaeogeography, Palaeoclimatology, Palaeoecology,179(3–4):173–188.doi:10.1016/S0031-0182(01)00413-8.
48.^ (英文)Maxwell, W. D.(1992年5月8日)."Permian and Early Triassic extinction of non-marine tetrapods".palaeontology,35:571–583.
49.^ Erwin DH(1990年5月8日).The End-Permian Mass Extinction.Annual Review of Ecology and Systematics,21:69–91.doi:10.1146/annurev.es.21.110190.000441.
50.^ 50.0 50.1 50.2 50.3 (英文)Knoll, A.H., Bambach, R.K., Payne, J.L., Pruss, S., and Fischer, W.W.(2009年5月8日).Paleophysiology and end-Permian mass extinction.Earth and Planetary Science Letters,256:295–313.於2008年7月4日查閱. Full contents may be available online at Paleophysiology and end-Permian mass extinction.於2008年7月4日查閱.
51.^ (英文)Lehrmann, D.J., Ramezan, J., Bowring, S.A., et al(2006年December月).Timing of recovery from the end-Permian extinction: Geochronologic and biostratigraphic constraints from south China.Geology,34:1053–1056.doi:10.1130/G22827A.1.
52.^ (英文)Sahney, S. and Benton, M.J.(2008年).Recovery from the most profound mass extinction of all time(PDF).Proceedings of the Royal Society: Biological,275:759.doi:10.1098/RSPB.2007.1370.
53.^ Ward PD, Montgomery DR, & Smith R(2000年).Altered river morphology in South Africa related to the Permian–Triassic extinction.Science,289(5485):1740–1743.doi:10.1126/science.289.5485.1740.
54.^ 54.0 54.1 (英文)Hallam A & Wignall PB(1997).Mass Extinctions and their Aftermath.Oxford University Press.ISBN 978-0198549161.
55^ (英文)Zi-qiang W(1996年).Recovery of vegetation from the terminal Permian mass extinction in North China.Review of Palaeobotany and Palynology,91:121–142.doi:10.1016/0034-6667(95)00069-0.
56^ (英文)Wagner PJ, Kosnik MA, & Lidgard S(2006年).Abundance Distributions imply Elevated Complexity of Post-Paleozoic Marine Ecosystems.Science,314(5803):1289–1292.doi:10.1126/science.1133795.
57^ (英文)Clapham, M.E., Bottjer, D.J. and Shen, S.(2006年).Decoupled diversity and ecology during the end-Guadalupian extinction (late Permian).Geological Society of America Abstracts with Programs,38(7):117.於2008年3月28日查閱.
58^ (英文)Foote, M.(1999年).Morphological diversity in the evolutionary radiation of Paleozoic and post-Paleozoic crinoids(PDF).Paleobiology,25(sp1):1–116.doi:[[doi:10.1666%2F0094-8373%281999%2925%5B1%3AMDITER%5D2.0.CO%3B2|10.1666/0094-8373(1999)25[1:MDITER]2.0.CO;2]].於2008年5月12日查閱.
59^ DOI:10.1146/annurev.earth.36.031207.124116
本引用來源將會在數分鐘後自動完成。您可以插隊或手動擴充
60.^ (英文)Benton, M.J.(2004).Vertebrate Paleontology.Blackwell Publishers,xii-452.ISBN 0-632-05614-2.
61.^ (英文)Ruben, J.A., and Jones, T.D.(2009年5月8日).Selective Factors Associated with the Origin of Fur and Feathers.American zoologist,40(4):585–596.doi:10.1093/icb/40.4.585.
62.^ (英文)Yates AM & Warren AA(2009年5月8日).The phylogeny of the 'higher' temnospondyls (Vertebrata: Choanata) and its implications for the monophyly and origins of the Stereospondyli.Zoological Journal of the Linnean Society,128(1):77-121.於2008年1月18日查閱.
63.^ (英文)Retallack GJ, Seyedolali A, Krull ES, Holser WT, Ambers CP, Kyte FT(1998年).Search for evidence of impact at the Permian–Triassic boundary in Antarctica and Australia.Geology,26(11):979–982.
64.^ 64.0 64.1 (英文)Becker L, Poreda RJ, Basu AR, Pope KO, Harrison TM, Nicholson C, Iasky R(2004年).Bedout: a possible end-Permian impact crater offshore of northwestern Australia.Science,304(5676):1469–1476.doi:10.1126/science.1093925.
65.^ (英文)Becker L, Poreda RJ, Hunt AG, Bunch TE, Rampino M(2001年).Impact event at the Permian–Triassic boundary: Evidence from extraterrestrial noble gases in fullerenes.Science,291(5508):1530–1533.doi:10.1126/science.1057243.
66.^ (英文)Basu AR, Petaev MI, Poreda RJ, Jacobsen SB, Becker L(2003年).Chondritic meteorite fragments associated with the Permian–Triassic boundary in Antarctica.Science,302(5649):1388–1392.doi:10.1126/science.1090852.
67.^ (英文)Kaiho K, Kajiwara Y, Nakano T, Miura Y, Kawahata H, Tazaki K, Ueshima M, Chen Z, Shi GR(2001年).End-Permian catastrophe by a bolide impact: Evidence of a gigantic release of sulfur from the mantle.Geology,29(9):815–818.於2007年10月22日查閱.
68.^ (英文)Farley KA, Mukhopadhyay S, Isozaki Y, Becker L, Poreda RJ(2001年).An extraterrestrial impact at the Permian–Triassic boundary?.Science,293(5539):2343.doi:10.1126/science.293.5539.2343a.
69.^ (英文)Koeberl C, Gilmour I, Reimold WU, Philippe Claeys P, Ivanov B(2002年).End-Permian catastrophe by bolide impact: Evidence of a gigantic release of sulfur from the mantle: Comment and Reply.Geology,30(9):855–856.doi:[[doi:10.1130%2F0091-7613%282002%29030%253C0855%3AEPCBBI%253E2.0.CO%3B2|10.1130/0091-7613(2002)030%3C0855:EPCBBI%3E2.0.CO;2]].
70.^ (英文)Isbell JL, Askin RA, Retallack GR(1999年).Search for evidence of impact at the Permian–Triassic boundary in Antarctica and Australia; discussion and reply.Geology,27(9):859–860.doi:[[doi:10.1130%2F0091-7613%281999%29027%253C0859%3ASFEOIA%253E2.3.CO%3B2|10.1130/0091-7613(1999)027%3C0859:SFEOIA%3E2.3.CO;2]].
71.^ (英文)Koeberl K, Farley KA, Peucker-Ehrenbrink B, Sephton MA(2004年).Geochemistry of the end-Permian extinction event in Austria and Italy: No evidence for an extraterrestrial component.Geology,32(12):1053–1056.doi:10.1130/G20907.1.
72.^ (英文)Langenhorst F, Kyte FT & Retallack GJ (2005). "Reexamination of quartz grains from the Permian–Triassic boundary section at Graphite Peak, Antarctica" (PDF). Lunar and Planetary Science Conference XXXVI. 於2007-07-13查閱.
73.^ (英文)von Frese RR, Potts L, Gaya-Pique L, Golynsky AV, Hernandez O, Kim J, Kim H & Hwang J(2006年).Abstract Permian–Triassic mascon in Antarctica.Eos Trans. AGU, Jt. Assem. Suppl.,87(36):Abstract T41A-08.於2007年10月22日查閱.
74.^ (英文)Von Frese, R.R.B.; L. V. Potts, S. B. Wells, T. E. Leftwich, H. R. Kim, J. W. Kim, A. V. Golynsky, O. Hernandez, and L. R. Gaya-Piqué(2009年).GRACE gravity evidence for an impact basin in Wilkes Land, Antarctica.Geochem. Geophys. Geosyst.,10:Q02014.doi:10.1029/2008GC002149.
75.^ (英文)Jones AP, Price GD, Price NJ, DeCarli PS, Clegg RA(2002年).Impact induced melting and the development of large igneous provinces.Earth and Planetary Science Letters,202(3):551–561.doi:10.1016/S0012-821X(02)00824-5.
76.^ 76.0 76.1 76.2 76.3 76.4 (英文)White RV(2002年).Earth’s biggest 'whodunnit': unravelling the clues in the case of the end-Permian mass extinction(PDF).Phil. Trans. Royal Society of London,360:2963-2985.doi:10.1098/rsta.2002.1097.於2008年1月12日查閱.
77.^ (英文)AHager, Bradford H (2001). "Giant Impact Craters Lead To Flood Basalts: A Viable Model". CCNet 33/2001: Abstract 50470.
78.^ (英文)Hagstrum, Jonathan T (2001). "Large Oceanic Impacts As The Cause Of Antipodal Hotspots And Global Mass Extinctions". CCNet 33/2001: Abstract 50288.
79.^ (英文)Zhou, M-F., Malpas, J, Song, X-Y, Robinson, PT, Sun, M, Kennedy, AK, Lesher, CM & Keays, RR(2002年).A temporal link between the Emeishan large igneous province (SW China) and the end-Guadalupian mass extinction.Earth and Planetary Science Letters,196(3–4):113–122.doi:10.1016/S0012-821X(01)00608-2.
80.^ (英文)Mundil, R., Ludwig, K.R., Metcalfe, I. & Renne, P.R(2004年).Age and Timing of the Permian Mass Extinctions: U/Pb Dating of Closed-System Zircons.Science,305(5691):1760–1763.doi:10.1126/science.1101012.
81.^ (英文)"Permian–Triassic Extinction - Volcanism"
82.^ (英文)Palfy J, Demeny A, Haas J, Htenyi M, Orchard MJ, & Veto I(2001年).Carbon isotope anomaly at the Triassic– Jurassic boundary from a marine section in Hungary.Geology,29(11):1047–1050.doi:[[doi:10.1130%2F0091-7613%282001%29029%253C1047%3ACIAAOG%253E2.0.CO%3B2|10.1130/0091-7613(2001)029%3C1047:CIAAOG%3E2.0.CO;2]].
83.^ 83.0 83.1 83.2 (英文)Berner, R.A.(2002年).Examination of hypotheses for the Permo-Triassic boundary extinction by carbon cycle modeling.Proceedings of the National Academy of Sciences,99:32095199.doi:10.1073/pnas.032095199.PMID 11917102.
84.^ 84.0 84.1 (英文)Dickens GR, O'Neil JR, Rea DK & Owen RM(1995年).Dissociation of oceanic methane hydrate as a cause of the carbon isotope excursion at the end of the Paleocene.Paleoceanography,10(6):965–71.doi:10.1029/95PA02087.
85.^ (英文)Schrag, D.P., Berner, R.A., Hoffman, P.F., and Halverson, G.P.(2002年).On the initiation of a snowball Earth.Geochemistry Geophysics Geosystems,3(6 doi=10.1029/2001GC000219):1036. Preliminary abstract at Schrag, D.P.(2001年June月).On the initiation of a snowball Earth.Geological Society of America.
86.^ 86.0 86.1 (英文)Benton, M.J.; Twitchett, R.J.(2003年).How to kill (almost) all life: the end-Permian extinction event.Trends in Ecology & Evolution,18(7):358–365.doi:10.1016/S0169-5347(03)00093-4.
87.^ (英文)Dickens GR(2001年).The potential volume of oceanic methane hydrates with variable external conditions.Organic Geochemistry,32(10):1179–1193.doi:10.1016/S0146-6380(01)00086-9.
88.^ (英文)Reichow MK, Saunders AD, White RV, Pringle MS, Al'Muhkhamedov AI, Medvedev AI & Kirda NP(2002年).40Ar/39Ar Dates from the West Siberian Basin: Siberian Flood Basalt Province Doubled.Science,296(5574):1846–1849.doi:10.1126/science.1071671.
89.^ (英文)Holser WT, Schoenlaub H-P, Attrep Jr M, Boeckelmann K, Klein P, Magaritz M, Orth CJ, Fenninger A, Jenny C, Kralik M, Mauritsch H, Pak E, Schramm J-F, Stattegger K & Schmoeller R(1989年).A unique geochemical record at the Permian/Triassic boundary.Nature,337(6202):39–44.doi:10.1038/337039a0.
90.^ (英文)Dobruskina IA(1987年).Phytogeography of Eurasia during the early Triassic.Palaeogeography, Palaeoclimatology, Palaeoecology,58(1-2):75–86.doi:10.1016/0031-0182(87)90007-1.
91.^ (英文)Newell ND(1971年).An Outline History of Tropical Organic Reefs(PDF).American Museum novitates,2465:1-37.於2007年11月3日查閱.
92.^ (英文)McRoberts, C.A., Furrer, H., Jones, D.S.(1997年).Palaeoenvironmental interpretation of a Triassic– Jurassic boundary section from western Austria based on palaeoecological and geochemical data..Palaeogeography Palaeoclimatology Palaeoecology,136(1-4):79– 95.doi:10.1016/S0031-0182(97)00074-6.
93.^ (英文)Wignall PB & Twitchett RJ(2002年).Permian–Triassic sedimentology of Jameson Land, East Greenland: Incised submarine channels in an anoxic basin.Journal of the Geological Society,159(6):691-703.doi:110.1144/0016-764900-120.
94.^ 94.0 94.1 (英文)Monastersky, R. (May 25, 1996), "Oxygen starvation decimated Permian oceans", Science News
95.^ 95.0 95.1 Zhang R, Follows, MJ, Grotzinger, JP, & Marshall J(2001年).Could the Late Permian deep ocean have been anoxic?.Paleoceanography,16(3):317–329.doi:10.1029/2000PA000522.
96.^ (英文)Kump LR, Pavlov A, & Arthur MA(2005年).Massive release of hydrogen sulfide to the surface ocean and atmosphere during intervals of oceanic anoxia.Geology,33(5):397–400.doi:10.1130/G21295.1.
97.^ (英文)Grice K, Cao C, Love GD, Bottcher ME, Twitchett RJ, Grosjean E, Summons RE, Turgeon SC, Dunning W & Yugan J(2005年).Photic Zone Euxinia During the Permian–Triassic Superanoxic Event.Science,307(5710):706-709.doi:10.1126/science.110432.
98.^ (英文)The Permian - Palaeos
