 藍白細尾鷯鶯
藍白細尾鷯鶯藍白細尾鷯鶯(學名:Malurus leucopterus)是雀形目鳥類的一個物種,屬於細尾鷯鶯科中的細尾鷯鶯屬。它們生活在澳大利亞中心的乾燥地帶;從昆士蘭州中心與南澳大利亞州到西澳大利亞州都有分布。如同其它細尾鷯鶯一樣,此物種也有顯著的兩性異形,在繁殖期,一個群居集體中的一隻或更多雄性會長出色彩明亮的羽毛。雌性全身為沙褐色,尾部羽毛淺藍 ;雌性體型比雄性小,繁殖期時雄性身體的羽毛為亮藍色,喙為黑色,翅膀為藍色。較年輕的性成熟雄性外形幾乎與雌性相同,通常這些雄性正在繁殖。一隊藍白細尾鷯鶯在春夏兩季會擁有一隻較大的、色彩明亮的雄性,這隻鳥由許多不顯眼的同樣是雄性的棕色小鳥陪伴著。得到承認的亞種共有三個;除開大陸上的亞種之外,一個亞種是在德克哈托格島上找到的,而另一個棲息於西澳大利亞州海岸外的巴羅島上。這些島嶼上的雄性在繁殖期羽毛是黑色而不是藍色。
特徵
 藍白細尾鷯鶯
藍白細尾鷯鶯藍白細尾鷯鶯長度為11至13.5厘米,是細尾鷯鶯屬中兩個體型最小的物種之一。雄性體重通常在7.2克至10.9克之間,而雌性體重通常在6.8克至11克之間。雄性的喙平均8.5毫米,雌性平均8.4毫米。喙相對較長、窄和尖,根基部較寬;在形狀上,它們和那些以探測或在附近捕捉昆蟲來取食的其他鳥類相似,喙的寬度比深度更長。而且此物種的喙比其他細尾鷯鶯的喙更細更尖。
完全成熟的成年鳥只是兩性異形的,雄性體型較大,色彩也與雌性不同。成年雌性為沙褐色,尾部淺藍, 喙為帶粉紅的暗黃色。擁有婚羽的雄性喙為黑色,翅膀與肩為白色,全身為鈷藍色或黑色(取決於亞種種類)。這些對比鮮明的白色羽毛在飛行和繁殖期的表演中尤其顯眼。有著蝕羽的雄性類似於雌性,但可以依靠其較黑的喙區分。兩性都擁有細長、獨特的尾羽,由它們身上一個向上的突出部分托舉著。尾部大約為6.25厘米長,尾部羽毛上有一條白色條紋,隱藏在身體的羽毛中。
雛鳥、剛會飛的小鳥和少年鳥只的羽毛為褐色,喙為粉褐色,尾羽比成年短。年輕雄性在晚夏或秋天結束前(在春夏的繁殖期之後)長出藍色的尾羽和較黑的喙,這時雌性長出淺藍色的尾羽。在下一個春天,所有雄性都會有強盛的繁殖力,並有成熟的泄殖腔突起,以儲存精細胞。與此相反,在繁殖期,繁殖力強的雌性會生長出一個水腫的抱卵點,是腹部上一個裸露的部分。進入第二年或第三年的雄性可能會在繁殖期長出有斑點的藍白色羽毛。在它們進入第四年之前,雄性已經得到了婚羽,此時肩羽、次級翼覆羽和次級飛羽為白色而身體其他部分為明亮的鈷藍色。所有性成熟的雄性都一年換羽兩次,一次是在冬或春季的繁殖期之前,另一次是在之後的秋天;雄性可能會很少見地直接從婚羽換羽至婚羽。由於鳥羽枝的表面平坦而扭曲,繁殖期雄性的藍色羽毛,尤其是耳覆羽十分閃亮。藍色羽毛也強烈地反射紫外線,所以可能對其他細尾鷯鶯來說較為耀眼,因為它們的色視覺達到了電磁波譜的此部分。
發聲
在1980年,泰德曼表現了Malurus leucopterus leuconotus亞種相互之間五種不同模式的鳴叫的特徵;而後普魯特和瓊斯認可島嶼亞種M. l. edouardi互相之間的鳴叫也是如此。最主要的一種鳴叫是一種囀鳴,兩性都會發出以建立領土和統一群體。這種鳴叫較長,是一種“升降鳴聲”,首先由3至5次鳴叫發出信號。雖然聲音看似微弱,但它會在矮小的灌叢帶上方傳播很長的距離。一種刺耳的“唧”聲常用於建立聯絡(尤其是母親與孩子之間的聯絡)和發出警告;其特徵表現為一系列“大聲而又突然”的鳴叫,頻率和強度不斷變化。成年會使用一種聲調高的吱吱聲來和更為遙遠的鳥只聯絡,這種吱吱聲可能間歇地和囀鳴混雜在一起。鳥巢周圍的雛鳥、剛會飛的小鳥和雌性會發出一種安靜、聲調高、短暫的嘰嘰聲。當成熟的雌性使用時,這種聲音和刺耳的鳴聲混雜在一起。當雛鳥正在被餵食時,它們也可能會發出“咕咕”的噪聲。副職的輔助者與餵食者也會發出這種聲音。
分布及棲息地
 藍白細尾鷯鶯
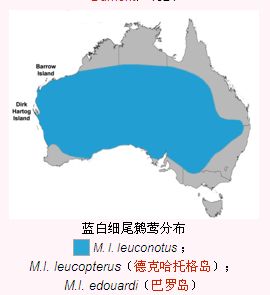
藍白細尾鷯鶯藍白細尾鷯鶯十分適應乾燥環境,M. l. leuconotus亞種遍及澳大利亞大陸南緯19至32度之間的乾旱和半乾旱環境。分布地占據了從黑德蘭港附近到南部的珀斯的澳大利亞西海岸,向東延伸至昆士蘭州的伊薩山,並沿著大分水嶺西部通過昆士蘭州中心和新南威爾斯的中西部,進入了維多利亞州的西北角和艾爾半島並穿過納拉伯平原.它們通常與其他種的細尾鷯鶯共同生活,包括雜色細尾鷯鶯(M. lamberti assimilis亞種)。藍白細尾鷯鶯常常棲息於石南荒原樹木較少的灌叢帶中,這些地方生長的主要是濱藜和藍澳藜屬的小灌木,或是三齒稃草和禾木等草類;它們同樣棲息於生長著竹節蓼的漫灘區域。M. l. leucopterus亞種棲息於德克哈托格島上相似的地方,而M. l. edouardi亞種也同樣如此。藍白細尾鷯鶯在其棲息地北部被紅背細尾鷯鶯取代了。
分類及命名
藍白細尾鷯鶯的一個標本最先於1818年9月被法國博物學家讓·雷內·康斯坦特·貴和約瑟夫· 保羅·蓋馬德收藏,它放在路易·德·弗雷西內環繞南半球航海的船上。此標本於一次海難中遺失,但一幅由傑奎斯·阿拉戈畫的名為Mérion leucoptère的畫得以倖存;據此,於1824年,法國鳥類學家查爾斯·杜孟特·德·聖克魯克斯對這種鳥進行了說明。此物種的學名源於古希臘語“leuko-”意思是“白色”而“pteron”意思是“翅膀”(中文名與學名意義不同)。
諷刺的是,最初的標本是黑色羽毛的來自德克哈托格島的亞種,此亞種在此後80年都沒有再發現過。與此同時,分布廣的藍色羽毛的亞種已被發現了,卻被約翰·古爾德在1865年描述為兩個不同的物種。他將來自來自內陸的新南威爾斯的標本叫做“白翅壯麗鶯”(White-winged Superb Warbler),“學名”M. cyanotus,而另一隻背和翅膀為白色,被說成是“白背壯麗鶯”(White-backed Superb Warbler),“學名”M. leuconotus。直到20世紀早期,這兩個藍羽毛的大陸的類型才被發現是同一個物種。 維多利亞州國家博物館的鳥類學家喬治·馬克在他1934年對該屬的修正中優先考慮了種名leuconotus,而更近代研究也紛紛效仿了此舉。此鳥的肩部之間的背部區域其實是裸露的,背部花紋為肩部上長出的羽毛(肩羽)向內部掃去形成的。這種變化使早期的博物學家產生錯誤,將白背和藍背的兩隻鳥說成是兩個物種。
藍白細尾鷯鶯常被認為是藍白鷦鷯,早期的觀察員,如維多利亞州的諾曼·法華羅洛,就曾用此名稱呼它們。然而和其它細尾鷯鶯一樣,藍白細尾鷯鶯和真正的鷦鷯科沒有親緣關係。從前它被分在鶲科中,後來又變成了鶯科的一員,直到1975年被分在新認可的細尾鷯鶯科中。在近代,DNA分析顯示細尾鷯鶯科與鴉總科中的吸蜜鳥科和斑啄果鳥科親緣關係較近。
藍白細尾鷯鶯是細尾鷯鶯科細尾鷯鶯屬的12個物種之一。它和澳大利亞的紅背細尾鷯鶯親緣關係最近,這兩個物種和黑白細尾鷯鶯一起組成了紐幾內亞的一個種系發生演化支,親緣關係為第二密切。這三個物種沒有頭部的裝飾和耳部的一束羽毛,全身黑色或藍色的羽毛十分均勻,肩部或翅膀搭配著對比強烈的顏色,因此它們外形顯著,並被鳥類學家理察·斯科德稱為“雙色鷦鷯”;它們在地理上互相填補空缺,遍及澳大利亞北部和紐幾內亞。
亞種
 藍白細尾鷯鶯
藍白細尾鷯鶯藍白細尾鷯鶯有三個被認可的亞種:
M. l. leuconotus
為澳大利亞大陸所特有,和其他亞種不同,只有此亞種的交配期雄性會展示顯眼的藍白色羽毛[18]此亞種的名字源於古希臘語,“leukos”意思是“白色”而“notos”意思是“背部”[5]其分布區南部的鳥往往比北方的體型小。[19]
M. l. leucopterus
只分布於澳大利亞西海岸外的德克哈托格島,交配期的雄性展示的是黑白色的羽毛。此亞種是三個中體型最小的,但尾羽長度占身體長度的比例較大。在德·弗雷西內的探險收集到種類標本的98年之後,它在1916年被湯姆·卡特再次收藏。
M. l. edouardi
與M. l. leucopterus相似,雄性也有黑白羽毛,它們只分布於巴羅島,巴羅島也在澳大利亞西海岸外。此亞種的鳥比M. l. leucopterus大,但尾羽較短。雌性的羽毛比起其他兩個亞種的灰褐色,較為偏淺紅褐色。A.J.坎貝爾在1901年說明了以上訊息。
M. l. leucopterus和M. l. edouardi通常體型都比大陸的亞種小,兩個亞種的家庭往往也都比大陸上那些由一雄性一雌性和一隻臨時的輔助者鳥構成的的家庭規模小。島嶼的種類和大陸的種類具有相似的社會結構,但島嶼上的繁殖配對的一窩卵的規模平均較小,孵化時間平均較長,而且存活的小鳥平均較少。此外,M. l. leuconotus由於出現範圍廣,被世界自然保護聯盟評級為無危 ,而島嶼的亞種由於脆弱的築巢地址易被人類的建設和居住所打擾,因此被澳大利亞政府評級為易危
生態與行為
 藍白細尾鷯鶯
藍白細尾鷯鶯藍白細尾鷯鶯移動方式通常為齊足跳,兩足都同時離開地面和著陸。但在表演“齧齒動物式跑步(rodent-run)”時也可能會跑動。身體的平衡是由比例較大的尾羽保持的,尾巴通常挺直,很少有靜止的時候。短而豐滿的翅膀為最初優秀的升空提供了條件,雖然對較長的路途用處不大,但對短距離飛行十分有用。
藍白細尾鷯鶯生活在複雜的社會群體中。家族由2至4隻鳥構成,通常有一隻棕色的,有時是藍色的雄性和一隻繁殖期的雌性。輔助者是前幾年撫養的鳥,在羽毛長出後它們會留在家庭中幫著撫養小鳥;它們可能是保留著棕色羽毛的雄性或是雌性。一個群體的鳥巢中的鳥並排在稠密的表面上互相用喙理羽毛。幾個小群體生活在一片領地內,組成一個家族,由一隻有著婚羽的藍色(或黑色)雄性負責管理。在藍色雄性在其家族內對其他棕色和部分藍色的雄性統治時,他只和一隻雌性一同巢居,也只撫養這隻雌性的後代。他是否會成為它領土內其它鳥巢的後代的父親尚不清楚。
每個家族有一篇指定的土地區域,所有成員都在此搜尋糧食與防禦。通常領土面積為4至6公頃,大小與區域內的雨水和資源的豐富程度有關;資源和昆蟲豐富的地區會出現較小的領土。此外,覓食領域在冬季更大,那時這些鳥花大量的時間於整個家族一起覓食。藍白細尾鷯鶯會比其他細尾鷯鶯占據更大的領土。
在對此物種的觀察中,“振翼”動作會以下兩種不同的情形中看到:第一種是雌性對雄性求偶表演的回應,可能是表達默認;第二種是少年鳥只請求食物,此信息由幫手傳達給較大的鳥只,未成熟的雄性傳達給年長的鳥。作此動作時細尾鷯鶯放低頭和尾,伸展並抖動其翅膀,安靜地將嘴保持張開。
雄性和雌性的成年藍白細尾鷯鶯都會運用一種“齧齒動物式跑步(rodent-run)”動作來使捕食者注意力離開小鳥的巢穴。此時頭、頸和尾羽放低,翅膀伸出,迅速的跑動將羽毛抖松,並發出一種持續的警報鳴聲。
食物
藍白細尾鷯鶯主要以昆蟲為食;其食物包括較為小型的甲蟲、臭蟲、蛾、螳螂、毛蟲以及更小的昆蟲如蜘蛛。較大的昆蟲通常由正在繁殖的雌性與其輔助者餵給雛鳥,輔助者中包括正在繁殖的雄性。成年與少年會從灌叢帶地面上跳過來搜尋食物,它們可能會用種子、濱藜或藜的果實以及聖彼得草的幼芽來補充飲食。在春夏兩季,鳥會在白天活躍地狂歡,唱著歌呆在糧草旁邊。此時的昆蟲數量眾多且易於捕捉,這使鳥在突擊捕捉食物之間的時間可以休息。白天高溫時一個群體常常一起躲避與休息。冬天食物較難找到,因此白天它們不得不持續不斷地尋找食物。
求偶與繁殖
細尾鷯鶯顯示出的偶外交配發生率為最高的物種之一,許多幼鳥是被非生身父親的雄性培養的。然而藍白細尾鷯鶯之間的求偶方法尚不清楚。藍色羽毛的雄性已好幾次被發現在其領土外搬運粉紅或紫色花瓣,這種花瓣在其他物種之間會起到向附近的雌性推薦雄性的作用。與此不同,巴羅島與德克哈托格島上的黑色羽毛雄性常常搬運的是藍色花瓣。花瓣的當搬運在家族領土外很遠處時,暗示著與其他雌性的交配將會發生,這還需要進一步的基因分析。
在另一種求偶表演中,雄性會面向雌性向前深深的鞠躬,喙將達到地面,並將羽毛伸展、平整,靠近地平線,持續最多20秒。做這個姿勢時,白色羽毛組成一條顯著的白色帶穿過深色的羽毛。
正在繁殖的雌性於春天開始築巢,建造的是半球形的建築物,由蜘蛛網,較好的草、 薊花冠毛和蔬菜冠毛構成,通常6至14厘米高,3至9毫米厚。每個巢都在一邊有一個小入口,它們通常被放置在靠近大地處的茂密灌木中。3至4個蛋的一窩卵在9月至1月通常到處都有放置,孵卵會持續大約14天。藍白細尾鷯鶯通常於春天在西澳大利亞州的西南部,但在澳大利亞北部和中部的乾旱區域則更加投機,在任何月份,一段降雨期之後幾乎都有繁殖記錄。孵卵由產子的雌性獨自完成,這時產子的雄性(棕色或藍色)和輔助者幫著餵食雛鳥與去除它們的糞囊。新孵化的小鳥是晚成雛的,孵化後立刻張嘴請求食物,生長柔和的羽毛系統,直到第三或第四天才睜開眼睛。雛鳥會在巢中停留10至11天,剛會飛的小鳥在離巢後還繼續需要3至4周的餵食。剛會飛的小鳥要么留著撫養下一窩鳥,要么移動到附近的區域去。一個配對在一個繁殖期內孵化並撫養兩巢鳥並不少見,繁殖雌性對輔助者的壓力往往會減少,餵養的總數目不會增加。同其他細尾鷯鶯一樣,藍白細尾鷯鶯特別易於被狹嘴金鵑巢寄生(Chalcites basalis)。被金鵑(C. lucidus)及黑耳金鵑(C. osculans)寄生的記錄很少。
進化歷史
島嶼的兩個亞種和大陸族群leuconotus亞種的遺傳距離都比和對方的更近;德克哈托格島距大陸2千米,而巴羅島距大陸56千米。族群間的基因流動在目前的間冰期開始的時候就已經存在了,有些是在8,000到10,000年前開始的,那時海平面更低,兩個島嶼都和大陸相連。
關於三種藍白細尾鷯鶯如何演化而來,共存在有三種理論。第一種認為祖先的情況為黑白色羽毛,隨著三種族群的分離,大陸的亞種演化出了藍白色羽毛。第二種假說認為在獨立的島嶼上集中演化出了黑白色羽毛。第三種認為祖先的藍白色羽毛的情況共同演化為了黑白色羽毛,後來大陸的種類重新演化出了藍色羽毛。
三個細尾鷯鶯物種的分布表明其祖先遍及紐幾內亞和澳大利亞北部,那時海平面較低,這兩個地區是由大陸橋連結著的。後來海平面上升,紐幾內亞的鳥演化成了黑白細尾鷯鶯,澳大利亞的種類成了紅背細尾鷯鶯和適應乾旱的藍白細尾鷯鶯;因此它們的族群分離開來。
