
基本信息
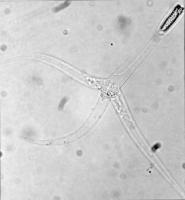 腦粘體蟲
腦粘體蟲分類學
腦粘體蟲Myxobolus cerebralis的分類學和命名及歸類到粘體動物亞門有著複雜的歷史。最初認為這種寄生蟲會感染魚腦(因此使用了種名“腦”cerebralis),但是很快就發在魚的神經系統發現了這一寄生蟲並發現其主要感染軟骨及骨骼。曾有人試圖將名稱改為能更準確描述這一生物的“軟骨粘體蟲Myxobolus chondrophagus”,但由於命名法的規則而無法更名。後來發現,先前稱為Triactinomyxon dubium和T. gyrosalmo (放射孢子綱Class Actinosporea)的生物事實上是腦粘體蟲的三角孢子蟲期,同時也將腦粘體蟲的生命周期擴展到三角孢子蟲期。同樣,其它放射孢子綱也被列入各種不同粘孢子蟲綱的生命周期。
今天,以前被認為是多細胞原生動物的粘體動物亞門也被許多科學家認為是動物,雖然它們的正式狀態並沒有改變。最近對分子領域的研究表明,這一類動物可能與兩側對稱動物或刺胞動物門有關。刺胞動物門可能在形態學上更接近,因為這兩類動物都有突出的極絲,但兩側對稱動物在一些遺傳學研究上可能更接近。
形態學
腦粘體蟲M. cerebralis有著許多不同分期,從單細胞到相對大的孢子,但並不是全部都得到詳細的研究。
三角孢子蟲期(Triactinomyxon stage)
這一階段稱為三角孢子蟲triactinomyxon spores,可以感染魚類,是一個約150微米(μm)長的單一類型。這一類型有三個突出或“尾巴”,每根“尾巴”的長度可達200微米。同時,這一類型的尾部有一個孢子原生質,孢子原生質含有64個生殖細胞並由細胞膜環繞。另外還有三個極囊,每個極囊都包含一個170~180 μm長的盤繞的極絲。這一階段和粘孢子蟲期的極絲快速射入宿主的體內,並創造一個供孢子原生質進入的通道。
孢子原生質期(Sporoplasm stage)
在接觸魚宿主並從極囊中發射出來後,孢子原生質被容納在三角孢子蟲進入上皮或腸道的中間“柱”。首先,孢子原生質經歷了有絲分裂來生成更多的變形蟲細胞。這些變形蟲細胞可以進入更深的組織層,以便能到達腦軟骨。
粘孢子蟲期(Myxosporean stage)
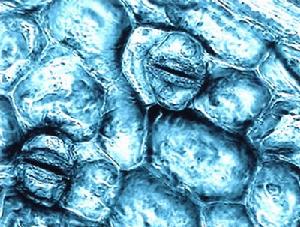 腦粘體蟲
腦粘體蟲通過魚體內的孢子增殖期演化而來的粘孢子蟲為晶體狀。這些粘孢子蟲的直徑約為10微米並由六個細胞組成。其中的兩個細胞形成極囊,兩個合併形成雙核的孢子原生質,另外兩個形成保護瓣。粘孢子蟲可以感染寡毛綱動物,並可在消化的魚軟骨殘餘中找到。由於不同屬之間形態學十分相似,一般很難將其與相關的品種區別開來。雖然腦粘體蟲是唯一可以在鮭亞目軟骨發現的粘孢子蟲,其它外觀相似的品種也可能在皮膚、神經系統和肌肉中存在。
生命周期
腦粘體蟲有一個雙宿主的生命周期,包括鮭魚和顫蚓科環節動物。到目前為止,唯一已知會感染腦粘體蟲的蠕蟲是正顫蚓Tubifex tubifex,雖然目前科學家表明T. tubifex可能會指幾個不同的品種。首先,粘孢子蟲被顫蚓科環節動物所消化。在蠕蟲的腸道內腔,孢子會排出極囊並用極絲附在腸道上皮。殼瓣沿縫脊打開,雙核生殖細胞則通過蠕蟲腸道上皮細胞之間滲透進入。細胞進行繁殖,並通過無性細胞二分裂的無卵核受精過程產生出很多變形蟲。在繁殖過程完成後,上皮細胞的胞間空隙超過了10個並排的蠕蟲並可能被感染。感染後約60~90天,寄生蟲經歷了有性細胞階段的孢子形成,並發展為泛孢子囊,其中每個都包含8個三角孢子蟲期的孢子。這些孢子由寡毛綱動物的肛門排放入水中。另外,魚類也可能因為吃到感染的寡毛綱動物而感染。已經感染的顫蚓會釋放三角孢子蟲至少1年。三角孢子蟲的孢子可以游在水中並通過皮膚感染鮭亞目動物。這些孢子進入魚體內只需要幾秒就已經足夠。只要五分鐘,生殖細胞稱為孢子原生質的液囊就可以進行魚的上皮細胞。而在一小時以內,孢子原生質就可以分裂成許多細胞在魚體內傳播。在魚的體內,這種繁殖包括細胞內和細胞外的軟骨內無性內生過程,也就是說新的細胞都從老細胞內生長而來。魚體內的最後階段是粘孢子蟲,粘孢子蟲是由孢子生殖形成。當魚體腐爛或被吃掉後,它們就進入環境中。最近的一些研究表明,一些魚在存活時會排出仍存活的粘孢子蟲。對於蠕蟲而言,粘孢子蟲是非常堅韌的:“腦粘體蟲的胞子可以在零下20°C存活至少3個月,13°C的土壤中存活5個月,並可以通過白斑狗魚Esox lucius或綠頭鴨Anas platyrhynchos的腸道而不失感染性”。三角孢子蟲存活時間更短,根據溫度的不同只能存活34天或更短時間。
病理學
腦粘體蟲對多種鮭亞目魚類的感染均有報導:其中有8種“大西洋”鮭亞目,斑鱒屬;4種“太平洋”鮭亞目,太平洋鮭屬;4種嘉魚,紅點鮭屬;茴魚,茴魚屬以及哲羅魚,哲羅鮭屬。腦粘體蟲會通過三角孢子蟲的附著和不同階段在組織、神經以及消化軟骨內的遷移對其魚宿主造成傷害。魚的尾巴會變黑,但除了軟骨的病變外,通常內臟器官看起來都較健康。其它症狀包括幼魚的骨骼變形及“鏇轉病”行為(追尾)。通常認為這些症狀是由於推動平衡導致,實際是由於脊髓和低位腦幹受損導致。試驗表明,魚可以在皮膚上就殺死粘體蟲(可能是抗生素的作用)。但是當它們進入中樞神經後,魚就無法再對其進行攻擊。不同品種之間的反應也並不相同。
 腦粘體蟲
腦粘體蟲易感性
魚的大小、年齡、三角孢子蟲的密度及水溫都會影響魚的感染率,而魚品種的影響目前還不大確定。由於5月齡以下的魚骨骼還未完全成骨,這一階段的魚受到該疾病的影響也最大。這會導致幼魚更可能感染並變形並提供更多軟骨來飼餵腦粘體蟲。一個對7個品種不同品系的研究發現暴露兩個小時後,溪紅點鮭和虹鱒(一個品系除外)比其它品種更容易感染腦粘體蟲,而公牛鱒、大鱗大麻哈魚、河鱒和北極茴魚則最不容易受到感染。但河鱒可能會攜帶這種寄生蟲。它們並不會表現出任何症狀但卻可能是腦粘體蟲的原始宿主。由於河鱒並不會表現出症狀,所以直到外來的虹鱒被引進到歐洲才發現了這一寄生蟲的存在。不同鮭亞目的易感性都在鮭亞目對鏇轉病的易感性中列出。
診斷
魚類對於鏇轉病中度或嚴重的臨床感染,可以根據初次感染35-80天后的行為及外觀變化進行推理診斷。由於受傷及日糧中缺乏色氨酸和抗壞血酸也可以導致類似的情況,最終診斷應當以在魚軟骨內發現粘孢子蟲為準。在嚴重感染的情況下,應當對軟骨進行顯微檢查並發現粘孢子蟲。在輕微感染的情況下,更常見的檢測是在查找粘孢子蟲前調查頭軟骨中胃蛋白酶和胰蛋白酶的消化情況(the pepsin-trypsin digest, PTD)。頭部和其它組織可以用組織病理學做進一步檢查並確認粘孢子蟲的位置和形態是否符合腦粘體蟲的特徵。組織部分的胞子血清學鑑定也可以使用抗孢子的抗生素。寄生蟲的鑑定可以用聚合酶鏈鎖反應(PCR)對腦粘體蟲415鹼基對上18S rRNA基因進行擴增並確認。初篩用的魚應當處於最可能感染該寄生蟲的生命階段。已經暴發該寄生蟲的國家都會定期使用這些技術進行檢測,而一些國家(像澳大利亞和加拿大)並沒有發生。
影響
雖然最初在中歐的河鱒(Salmo trutta)和東北亞的其它鮭魚上發現了野生病原,虹鱒(Oncorhynchus mykiss)對病原的傳播卻大大增加了這種寄生蟲的影響。由於無法對腦粘體蟲產生自然免疫,虹鱒特別易感並可以傳播很多孢子給同地區的其它魚類甚至是抗病品種,如河鱒,並因此攜帶過多寄生蟲並導致大量死亡。在腦粘體蟲暴發的地區,可能導致魚群的大量減少甚至滅絕。
在歐洲的影響
 腦粘體蟲
腦粘體蟲由於這一物種已經成為歐洲的地方流行病並使魚群得到免疫,腦粘體蟲對歐洲的影響在某種程度上得到減輕。作為最易感的物種,虹鱒並非歐洲的本土魚類。由於野生繁殖的魚群很少,也很少有足夠小的野生虹鱒會感染這種寄生蟲。另一方面,它們廣泛養殖於游釣水域及該寄生蟲影響最大的水產業。歐洲設計用於防止虹鱒魚苗受到感染的孵化及培育方法也被證明是成功的。這些技術包括在無孢子的水域進行孵化並在水箱或水道中培育魚苗到成骨期。這些方法特別關注水源質量,以避免在換水時引入孢子。在骨骼成骨並被認為可在臨床上對此寄生蟲產生抵抗力後,魚苗才被移到地面池塘中。
在紐西蘭的影響
腦粘體蟲1971年首次在紐西蘭發現。這種寄生蟲只在遠離多數水產養殖區的南島的河流發現。另外,紐西蘭對鮭亞目的商業化養殖也很少感染鏇轉病,而且這種寄生蟲也並沒有感染這裡的本地鮭亞目品種。這種寄生蟲存在的一個間接重要影響是對出口到澳大利亞鮭魚產品的嚴格檢疫。
1956年,賓夕法尼亞州由於從歐洲引進了已經感染的鱒魚而將腦粘體蟲引入,並傳播至南部和西部地區。這也是腦粘體蟲在北美首次得到報導。直至1980年代,鏇轉病仍被認為是在孵化池虹鱒的管理問題所造成。但是最近在落基山脈諸州(科羅拉多州、懷俄明州、猶他州、蒙大納州、愛達荷州及新墨西哥州)的天然水域也存在了該寄生蟲,並在這裡的一些游釣河流造成了較高死亡率。美國西部的一些河流甚至損失了90%的鱒魚。另外,鏇轉病也影響了在旅遊業中相當重要的休閒遊釣,這一行業在美國西部一些州份的經濟中也占據了較大比重。例如,蒙大拿鏇轉病特別小組(Whirling Disease Task Force)估計鮭魚相關休閒娛樂產業產生的消費僅在蒙大納州就有300,000,000美元。。更糟的是,一些腦粘體蟲感染的魚類(公牛鱒、山鱒及硬頭鱒)已經受到威脅或瀕臨絕種。科羅拉多州和蒙大納州受到影響最為嚴重,而加利福尼亞州、密西根州和紐約州受到的影響則最輕,具體的原因仍不清楚,但可能與環境條件有關。
防控措施
一些生物學家試圖在三角孢子蟲成熟前將其滅活。在實驗室里,目前只有強酸或強鹼、中高濃度的鹽類或電可以提前讓極絲失活。而神經化學、刺細胞動物的化學致敏物質及鱒魚黏膜均無效,同時也會令魚受到麻醉或死亡。如果孢子能得到滅活則無法感染魚,但仍不清楚是否實驗室的方法能否用於野生魚類。一些魚的品系比其它魚更具抗病力,甚至是同一品種的。使用抗病品系可能會幫助減少水產養殖中鏇轉病的發病率及嚴重性。也有證明表明魚群可在一定時間後自行產生抗病力。另外,水產業也可以通過不使用地面池塘養殖幼魚來避免感染腦粘體蟲。這可以使幼魚不會感染到顫蚓並更容易通過過濾、氯化及紫外消毒來消滅孢子和寡毛綱動物。減少顫蚓群體的技術包括:定期對孵化池或養殖用池塘消毒,並在室內無病原的水中飼養小鱒魚。光滑表面的混凝土或保持清潔的塑膠水道及無污染的水可以保持養殖環境免受疾病影響。最後,一些藥物如呋喃唑酮(痢特靈)、苯菌靈、煙麴黴素、氯胍和氯胺酮可以阻止孢子發育並減少感染率。舉個例子,有個研究表明給虹鱒Oncorhynchus mykiss飼餵煙麴黴素可以將受到感染的魚總數從73%~100%減少至10%~20%。不幸的是,這種治療方法並不適用於野生鱒魚,而且在美國食品藥品監督管理局(FDA)批准進行的研究中也沒有發現任何藥物能有效治療這一疾病。游釣人員可以通過一些方法來防止這種寄生蟲的傳播。如在不同地方釣魚時應事先清潔釣具並不將魚從一個水域帶到一水域,這樣也能避免不同水道間的交叉污染。孢子可以隱藏在氈底的涉水鞋下,需要用10%氯漂白劑和水進行消毒至少15分鐘並徹底沖洗。由於胴體中的孢子會釋放到水中,魚骨或內臟不能暴露在任何水體中。鮭魚和鱒魚不能用作誘餌。
