
病理簡介
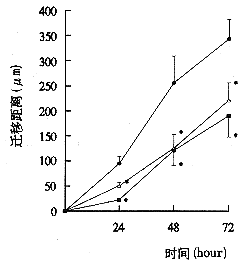 細胞遷移
細胞遷移 病理髮現
 解聚
解聚 雖然1939年科學家阿爾伯特•山特吉爾吉(A.Szent-Györgyi)就已發現細胞骨架的成分—肌動蛋白和肌球蛋白,但是因為電子顯微鏡製作樣本時需要對樣品進行0到4°C的低溫固定,在這樣的溫度下細胞骨架會被破壞,即所謂的“解聚”。所以當時認為細胞質不過是一“蛋白湯”,各種細胞器懸浮於細胞質液(Cytosol)中。
 細胞遷移
細胞遷移 但在60年代後,人們使用戊二醛常溫固定的方法開始逐漸發現細胞骨架。科學家發現,細胞骨架在這個細胞遷移過程起到承載和支撐的作用。在20世紀末21世紀初,科學家對細胞遷移複雜機理的認識有了非常大的進步,對細胞與基質的粘著,非對稱性極化和胞內分層運動都有了進一步的了解。但是整個過程其實仍未被了解透徹,很多中間過程就是連起作用的物質都未明。科學家對其中部分需要進行假設,再進一步通過實驗去證實。
研究技術
 細胞遷移
細胞遷移 新科技對細胞遷移研究起到了極大的推動作用。科學家通過ECIS技術(ElectricCell-substrateImpedanceSensing;電子細胞基質阻抗判斷)可以觀察到細胞在傳統細胞培養甚至是液體環境中的移動行為。根據ECIS技術觀測細胞電學參數的能力,ECIS技術還可以量化測量腫瘤細胞遷移過程中細胞層形態變化。同樣是在腫瘤研究領域,ATIM(Fluorescence-AssistedTransmigrationInvasionandMotilityAssay,螢光協助轉移侵入和運動分析法)提供了快速定量細胞侵入(細胞從一個區域進入另一區域)的更好方法,允許檢測大量樣品和不同條件下時間依賴性侵入。
更重要的是,這一系統可以方便地通過在多孔膜上增加胞外基質的厚度來監測細胞侵入結構的深度。韓國延世大學的朴宗哲和朴峰珠則發展出一套細胞跟蹤系統。它是由計算機輔助的時間流逝顯示微觀複製系統,其中有影象形成過程軟體,其程式編制含有自動影象分析和自設計CO2微小細胞培育器,它的功能是在一個倒置顯微鏡平台上,對於細胞遷移進行迅速而精確的分析,從而形成對於細胞的培育。目前已知他們運用這一計算機輔助系統計算了外細胞間質(ECMs)覆蓋表面的細胞遷移過程。
斑馬魚是目前在該領域最常用於研究的生物。細胞遷移是脊椎動物胚胎髮育的核心過程之一。細胞從原分裂生成的部位移動到目的部位就是細胞的遷移。斑馬魚有著很大的優勢,首先是其胚胎能在母體外發育,速度快,受精24小時後身體的器官已大部分就位。而且斑馬魚繁殖量大,容易對之進行變異。還有其胚胎透明,在高解析度快進攝影技術的幫助下,人們可以很好的觀察到細胞遷移的過程,還可以利用綠色螢光蛋白(GFP)可以觀察到細胞在斑馬魚體內的分布情況。
開關調節
 細胞遷移--開關調節
細胞遷移--開關調節 GEF和GAP如同兩隻手,一手打開開關,一手關閉開關。但是,這兩隻手必須協調工作,才能精確地調控細胞遷移。在過去的研究中,科學家們對“打開開關的手”研究甚多,而對“關閉開關的手”如何作用缺乏了解。朱學良研究員的小組發現一種名為Nudel的蛋白質能控制這隻“關閉開關的手”,在必要時能將其與開關隔離,從而保證足夠多的開關處於開啟的狀態。
事實上,Nudel通過與GAP結合,阻擋了GAP對Cdc42的作用。但如果Cdc42過量,也能通過與Nudel競爭結合GAP而失活。在實驗中,研究人員發現缺失了Nudel的細胞爬行受到了很大幹擾,在600分鐘的視頻中,正常細胞已經運動了很長距離,而缺失細胞則幾乎在原地一動沒動。這一研究揭示了一條新的調節細胞遷移開關的信號通路,對於深入了解細胞遷移的調節機制有重要意義。該研究也為認識相關疾病的機理提供了一條新線索。
參與遷移
細胞遷移需要內外因素的配合。外部的因素指的是細胞外的信號分子。內部因素則指細胞的信號傳導系統和執行運動的細胞骨架和分子馬達,還有參與粘著斑形成的各種分子(關於參與形成粘著斑的各種分子請見突出與底質的粘著)。細胞外信號結合胞膜受體完成其使命後,需要細胞內信號分子接力,將運動信息進一步傳給細胞遷移的執行單位——細胞骨架和分子馬達。種類繁多的細胞內信號分子會相互作用,影響後述這兩種分子的分布,結構和活性,達到精細調整細胞運動的目的。
信號分子
細胞外
 細胞遷移--細胞外
細胞遷移--細胞外 細胞內
胞外信號種類繁多,但是當它們與細胞膜上受體結合之後,細胞內起作用的途徑卻只有有限的幾種。而與細胞遷移有關的信號傳導過程如下:信號分子結合到膜上受體,或者是激活與受體偶聯的蛋白質—大G蛋白,或者先是激活受體酪氨酸激酶,再激活下游的小G蛋白Ras。G蛋白是一個很大的家族,包括Rho,Rac,Ras等小家族,它們在細胞中扮演著信號傳導開關的角色。當它們與GDP結合時,呈現失活狀態。在鳥嘌呤交換因子(英文:Guaninexchangefactor,簡稱GEF)的幫助下,G蛋白脫離GDP並與GTP結合,進入激活狀態。G蛋白的GTP會被GTP酶激活蛋白(英文GTPase-activatingproteins,簡稱GAP)水解,並釋放出其中的能量,讓G蛋白行使其功能。就是說,G蛋白通過這一GTP/GDP循環在激活/失活狀態中迴旋,傳遞信號。當G蛋白被激活後,它下游的多種分子會被激活。從插圖2中可見,這些下游分子本身會形成網路,相互制約,或者是相輔相成。它們調控著細胞遷移中各個方面。它們作用的詳細情況請見文章中的相應章節。而致癌物質也可以通過這些信號傳導通路發揮其負面作用,如強烈致癌物質佛波酯(Phorbolester)。佛波酯會不可逆地激活細胞的RasGRP3/4,以激活Ras,Ras會再激活蛋白激酶C(ProteinkinaseC,PKC)。後者是調節細胞分裂和分化的酶。它被佛波酯不正常的激活,有可能對癌症的產生起促進作用。研究還發現,佛波酯對黑素瘤(melanoma)細胞轉移到肺部有促進作用。而細菌者,如志賀氏菌會在宿主胞膜上打洞,向細胞質注入效應蛋白質,激活宿主Rac和Cdc42,調整細胞的微絲網路,以使自己順利進入宿主內。
細胞骨架
細胞骨架的定義分為狹義和廣義兩種,前者是微絲,微管和中間纖維的總稱,它們存在於細胞質內,又被稱為“胞質骨架”。後者還包括細胞外基質(extracellularmatrix),核骨架(nucleoskeleton)和核纖層(nuclearlamina)。細胞骨架是細胞內運動,細胞器固定,細胞外型維持,信號傳導和細胞分裂的物質基礎之一。
 細胞遷移
細胞遷移 微絲和其結合蛋白
微絲是由肌動蛋白(Actin)組成的直徑約為7nm纖維結構。肌動蛋白單體(又被稱為G-Actin,全稱為球狀肌動蛋白,GlobularActin,下文簡稱G肌動蛋白)為球形,其表面上有一ATP結合位點。肌動蛋白單體一個接一個連成一串肌動蛋白鏈,兩串這樣的肌動蛋白鏈互相纏繞扭曲成一股微絲。這種肌動蛋白多聚體又被稱為纖維形肌動蛋白(F-Actin,FibrousActin)。
微絲能被組裝和去組裝。當單體上結合的是ATP時,就會有較高的相互親和力,單體趨向於聚合成多聚體,就是組裝。而當ATP水解成ADP後,單體親和力就會下降,多聚體趨向解聚,即是去組裝。高ATP濃度有利於微絲的組裝。所以當將細胞質放入富含ATP的溶液時,細胞質會因為微絲的大量組裝迅速凝固成膠。而微絲的兩端組裝速度並不一樣。快的一端(+極)比慢的一端(-極)快上5到10倍。當ATP濃度達一定臨界值時,可以觀察到+極組裝而-極同時去組裝的現象,被命為“踏車”。微絲的組裝和去組裝受到細胞質內多種蛋白的調節,這些蛋白能結合到微絲上,影響其組裝去組裝速度,被稱之為微絲結合蛋白(associationprotein)。微絲的組裝先需要“核化”(nucleation),即幾個單體首先聚合,其它單體再與之結合成更大的多聚體。Arp複合體(Arp:Actinrelated-protein)是一種能與肌動蛋白結合的蛋白,它起到模板的作用,促進肌動蛋白的多聚化。Arp複合體由Arp2,Arp3和其它5種蛋白構成,也寫成Arp2/3複合體。封閉蛋白(end-blockingprotein)則是微絲兩端的“帽子”。
當這種蛋白結合到微絲上時,微絲的組裝和去組裝就會停止。這對一些長度固定的蛋白來說很重要,如細肌絲。而前纖維蛋白(Profilin,或譯G肌動蛋白結合蛋白)則是促進多聚的,相應地促解聚的蛋白則有絲切蛋白(Cofilin)。纖絲切割蛋白(filamentseveringprotein),如溶膠蛋白(Gelsolin),能將微絲從中間切斷。粘著斑蛋白(Vinculin)則能固定微絲到細胞膜上,形成粘著斑。交聯蛋白(cross-linkingprotein)有兩個以上肌動蛋白結合位點,起到連線微絲的作用,其中,絲束蛋白(fimbrin)幫助微絲結成束狀,而細絲蛋白(filamin)則將微絲交聯成網狀。
微管
微管是另一種具有極性的細胞骨架。它是由13條原纖維(protofilament)構成的中空管狀結構,直徑22—25nm。每一條原纖維由微管蛋白二聚體線性排列而成。微管蛋白二聚體由結構相似的α和β球蛋白構成,兩種亞基均可結合GTP,α球蛋白結合的GTP從不發生水解或交換,是α球蛋白的固有組成部分,β球白結合的GTP可發生水解,結合的GDP可交換為GTP,可見β亞基也是一種G蛋白。微管和微絲一樣,具有生長速度較+端和較慢的-端。微管在細胞內起支撐作用。另外它還是兩種運載分子:驅動蛋白(Kinesin)和發動蛋白(Dynein),的行走軌道。微管,可能連帶負在其上的發動蛋白會發放信號促進粘著斑的解聚,後者是粘著斑的周轉和尾部與底質分離過程中重要的一步。
中間纖維
中間纖維(intermediatefilaments,IF)直徑10nm左右,介於微絲和微管之間。與後兩者不同的是中間纖維是最穩定的細胞骨架成分,它主要起支撐作用。中間纖維在細胞中圍繞著細胞核分布,成束成網,並擴展到細胞質膜,與質膜相連結。中間纖維沒有正負極性。角蛋白是中間纖維中的一類,分子量約40~-0KD,出現在表皮細胞中,在人類上皮細胞中有20多種不同的角蛋白,分為α和β兩類。角蛋白賦予細胞體一定的剛性。癌細胞需要對角蛋白進行重新分布,以使自身變得柔韌,可以通過基底膜或血管壁上的細小孔洞。
分子馬達
 分子馬達
分子馬達 過程簡述
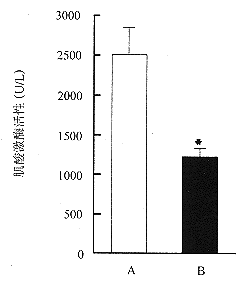 細胞遷移
細胞遷移 值得一提的是,細胞在前進的過程中,可以不斷改變其前進的方向。在顯微鏡下觀察大腸桿菌(Escherichiacoli)尋找食物時的運動,可見細胞先向前直線移動一段時間,然後會停下來並且調整一下方向,然後又再作直線移動。如此不斷反覆。可見細胞內調控能力的有效和精確。細胞極化後,細胞的前端會伸出極狀足(請見細胞前端突出)。極狀足伸出後,會與細胞前方的底質附著;粘著處會形成一種固定結構,名曰粘著斑(請見突出與底質的粘著)。此時,胞體主體會被牽拉向前(請見細胞體前移);最後細胞的後端與底質剝離(請見牽引尾部往前)。這樣前進的4個步驟完成,並準備下一次循環。不同的細胞,它們直線運動的速度和持續的時間是不同的。使用分子干擾技術可以很好的研究這兩者。一般來說,細胞直線前進速度越慢,其保持直線運動的時間就越長,例外是魚的上皮細胞,它能夠在快速遷移的同時,顯示出長時間保持直線運動的能力。
極化病理
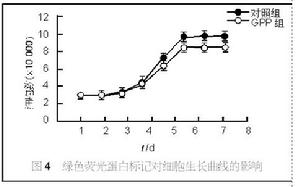
 細胞遷移
細胞遷移 這種mRNA的局部匯聚反過來卻需要微絲還有微管的協助。這種mRNA匯聚的發生對細胞遷移有極大的好處。一方面降低了細胞對之調控或降解的難度,另一方面它們的新生翻譯產物互相靠近又有利於複合體的快速組裝(這種現象被稱做共同翻譯組裝,英文:cotranslationalassembly),同時也保證了各自的正確摺疊,避免與其他細胞成分發生負面的相互作用。
鈣離子的分布與上面提到的不同。Ca2+前端分布濃度低,而尾端高。如果在一遷移中的白細胞側邊放入遷移信號物,人們發現細胞內的鈣離子水平先是總體升高,然後再分布成前低後高的狀態,細胞轉彎。如果此時撤銷新放入的信號,白細胞會接著按照新的信號物濃度繼續其遷移。很多微絲結合蛋白,如I和II型肌球蛋白,凝膠溶素,輔肌動蛋白(輔肌動蛋白)和絲束蛋白(fimbrin)都受到鈣離子的調控。因此鈣離子是細胞由溶膠變凝膠(sol-to-gel)過程中重要的一員。前端蓋離子濃度低,能激活I型肌球蛋白,抑制微絲分解蛋白,撥轉由鈣離子調控的微絲交聯蛋白,因此有利於微絲網路的形成。而尾部高鈣離子濃度則會導致微絲的解聚,激活凝溶膠蛋白,促成溶膠狀態的出現,而且會激活II型肌球蛋白,使外皮微絲網路收縮。總的說來,鈣離子的胞內濃度梯度對微絲的周轉起到了很重要的作用。
運動步驟
 細胞遷移
細胞遷移 其中一部分移行於外胚層下方,將來會分化為色素細胞,而那些行走的稍深一點的細胞,會形成後來交感神經的神經節細胞,腎上腺髓質。而頸部和骶部的神經嵴細胞則會沿著身體縱軸移到腸壁。就是說,日後的組織腸神經叢,神經節神經元,腎上腺的嗜鉻細胞(chromaffin)都是由神經嵴細胞遷移分化得出的。值得注意的是,在此過程中,沿途的不遷移細胞可能會影響遷移細胞的行為,改變它們的去向,甚至決定遷移細胞是否能存活。
同來自神經嵴的性細胞,血細胞前體和色素細胞都受到一種Kit—Steel因子機制的調節。Kit是一種跨膜受體,其配體是Steel因子。沿途的細胞或者是終點處的細胞會表達Steel因子,激活遷移經過的細胞上的Kit受體。而Kit受體的激活是這些細胞存活和增殖的前提。在一個個體中,兩者之中的任一者出現突變,患者的體色,血細胞供應和性細胞的形成都會出現異常,例如患者額頭可見一白斑。
相關詞條
參考資料
1.http://www.lunwentianxia.com/product.sf.3109144.1/
2.http://www.zaxue.com/article/4/2007/2007082925845.html
3.http://www.doctorsky.cn/medical-archive/other/20080531/6834.html
