基本特徵
概述
 病毒
病毒原指一種動物來源的毒素。“virus”一詞源於拉丁文。病毒能增殖、遺傳和演化,因而具有生命最基本的特徵,至今對它還沒有公認的定義。其主要特點是:
①形體極其微小,一般都能通過細菌濾器,因此病毒原叫“濾過性病毒”,必須在電子顯微鏡下才能觀察。
②沒有細胞構造,其主要成分僅為核酸和蛋白質兩種,故又稱“分子生物”;
③每一種病毒只含一種核酸,不是DNA就是RNA。
④既無產能酶系,也無蛋白質和核酸合成酶系,只能利用宿主活細胞內現成代謝系統合成自身的核酸和蛋白質成分。
⑤以核酸和蛋白質等“元件”的裝配實現其大量繁殖。
⑥在離體條件下,能以無生命的生物大分子狀態存在,並長期保持其侵染活力。
⑦對一般抗生素不敏感,但對干擾素敏感。
⑧有些病毒的核酸還能整合到宿主的基因組中,並誘發潛伏性感染。
簡單理解
病毒,是一類不具細胞結構,具有遺傳、複製等生命特徵的微生物。病毒同所有的生物一樣,具有遺傳、變異、進化的能力,是一種體積非常微小,結構極其簡單的生命形式,病毒有高度的寄生性,完全依賴宿主細胞的能量和代謝系統,獲取生命活動所需的物質和能量,離開宿主細胞,它只是一個大化學分子,停止活動,可製成蛋白質結晶,為一個非生命體,遇到宿主細胞它會通過吸附、進入、複製、裝配、釋放子代病毒而顯示典型的生命體特徵,所以病毒是介於生物與非生物的一種原始的生命體。
病毒的分類:
從遺傳物質分類:DNA病毒、RNA病毒、蛋白質病毒(如:朊病毒)
從病毒結構分類:真病毒(Euvirus,簡稱病毒)和亞病毒(Subvirus,包括類病毒、擬病毒、朊病毒)
從寄主類型分類:噬菌體(細菌病毒)、植物病毒(如菸草花葉病毒)、動物病毒(如禽流感病毒、天花病毒、HⅣ等)
從性質來分:溫和病毒(HⅣ)、烈性病毒(狂犬病毒)。
病毒的形態
⑴球狀病毒;⑵桿狀病毒;⑶磚形病毒;⑷冠狀病毒;⑸絲狀病毒
⑹鏈狀病毒;⑺有包膜的球狀病毒;⑻具有球狀頭部的病毒;⑼封於包含體內的昆蟲病毒。
病毒粒的對稱體制:
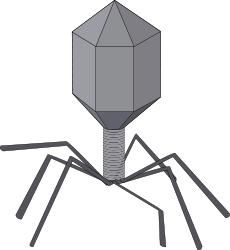
病毒粒的對稱體制只有兩種,即螺鏇對稱(代表菸草花葉病毒)和二十面體對稱(等軸對稱,代表腺病毒)。一些結構較複雜的病毒,實質上是上述兩種對稱相結合的結果,故稱作複合對稱(代表T偶數噬菌體)
病毒的大小
 病毒
病毒病毒的組成
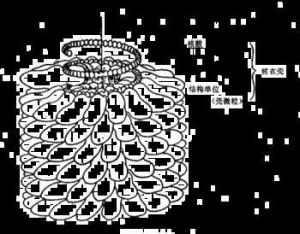
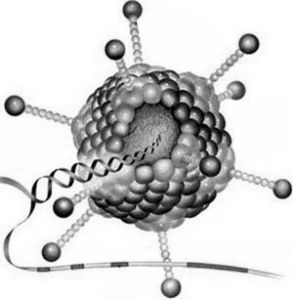
病毒主要由內部的遺傳物質和蛋白質外殼組成。由於病毒是一類非細胞生物體,故單個病毒個體不能稱作"單細胞",這樣就產生了病毒粒或病毒體(virion)。病毒粒有時也稱病毒顆粒或病毒粒子(virusparticle),專指成熟的、結構完整的和有感染性的單個病毒。核酸位於它的中心,稱為核心(core)或基因組(genome),蛋白質包圍在核心周圍,形成了衣殼(capsid).衣殼是病毒粒的主要支架結構和抗原成分,有保護核酸等作用。衣殼是由許多在電鏡下可辨別的形態學亞單位(subunit)——衣殼粒(capsomere)所構成。核心和衣殼合稱核心殼(nucleocapsid)。有些較複雜的病毒,(一般為動物病毒,如流感病毒),其核心殼外還被一層含蛋白質或糖蛋白(glycoprotein)的類脂雙層膜覆蓋著,這層膜稱為包膜(envelope)。包膜中的類脂來自宿主細胞膜。有的包膜上還長有刺突(spike)等附屬物。包膜的有無及其性質與該病毒的宿主專一性和侵入等功能有關。昆蟲病毒中有1類多角體病毒,其核殼被蛋白晶體所包被,形成多角形包涵體。
病毒的複製過程叫做複製周期。其大致可分為連續的五個階段:吸附、侵入、增殖、成熟(裝配)、裂解(釋放)。
病毒的結構
1.病毒的基本結構有核心和衣殼,二者形成核衣殼。核心位於病毒體的中心,為核酸,為病毒的複製、遺傳和變異提供遺傳信息;衣殼是包圍在核酸外面的蛋白質外殼。
衣殼的功能:①具有抗原性;②保護核酸;③介導病毒與宿主細胞結合。
2.病毒的輔助結構
有些病毒核衣殼外還有一層脂蛋白雙層膜狀結構,是病毒以出芽方式釋放,穿過宿主細胞膜或核膜時獲得的,稱之為包膜。在包膜表面有病毒編碼的糖蛋白,鑲嵌成釘狀突起,稱為刺突。有包膜病毒對有機溶劑敏感。
包膜功能:①保護核衣殼;②促進病毒與宿主細胞的吸附;③具有抗原性。
病毒出現假說:
1.蛋白質、核酸遺失說:
大生物(此處大生物意思是具有細胞結構的生物,區別於病毒的非細胞結構生物)由於細胞脫落和破裂,導致游離的蛋白質和DNA、RNA的出現,在某種情況下,這些蛋白質由於化學作用形成了一個內部可容納小分子的結構,裡面裹著DNA或者RNA,甚至單獨的蛋白質和單獨的DNA、RNA游離,這些散落的游離的分子,有一些個別的和大生物細胞膜有親和性,大生物細胞通過吞噬作用使其進入細胞,其DNA、RNA得以表達,然後通過進化形成成熟的病毒。
2.生命起源說:
病毒是最原始的生命體,早在沒有細胞之前就有病毒存在,那時的病毒還只限於蛋白質和核酸,沒有表現出病毒的寄生特徵,當細胞體生物出現之後,個別這種蛋白質和核酸或他們的複合體表現出寄生性,這些散落的游離的分子,有一些個別的和大生物細胞膜有親和性,大生物細胞通過吞噬作用使其進入細胞,其DNA、RNA得以表達,然後通過進化形成成熟的病毒。
來源
在病毒大家庭中,有一種病毒有著特殊的地位,這就是菸草花葉病毒。無論是病毒的發現,還是後來對病毒的深入研究,菸草花葉病毒都是病毒學工作者的主要研究對象,起著與眾不同的作用。發現
 病毒
病毒1892年,俄國的伊萬諾夫斯基(Ivanovski)重複了麥爾的試驗,證實了麥爾所看到的現象,而且進一步發現,患病菸草植株的葉片汁液,通過細菌過濾器後,還能引發健康的菸草植株發生花葉病。這種現象起碼可以說明,致病的病原體不是細菌,但伊萬諾夫斯基將其解釋為是由於細菌產生的毒素而引起。生活在巴斯德的細菌致病說的極盛時代,伊萬諾夫斯基未能做進一步的思考,從而錯失了一次獲得重大發現的機會。
1898年,荷蘭細菌學家貝傑林克(Beijerinck)同樣證實了麥爾的觀察結果,並同伊萬諾夫斯基一樣,發現菸草花葉病病原能夠通過細菌過濾器。但貝傑林克想得更深入。他把菸草花葉病株的汁液置於瓊脂凝膠塊的表面,發現感染菸草花葉病的物質在凝膠中以適度的速度擴散,而細菌仍滯留於瓊脂的表面。從這些實驗結果,貝傑林克指出,引起菸草花葉病的致病因子有三個特點:1,能通過細菌過濾器;2,僅能在感染的細胞內繁殖;3,在體外非生命物質中不能生長。根據這幾個特點他提出這種致病因子不是細菌,而是一種新的物質,稱為“有感染性的活的流質”,並取名為病毒,拉丁名叫“Virus”。
神奇的病毒“誕生”了!
幾乎是同時,德國細菌學家勒夫勒(Loeffler)和費羅施(Frosh)發現引起牛口蹄疫的病原也可以通過細菌濾器,從而再次證明伊萬諾夫斯基和貝傑林克的重大發現。
“virus”一詞源於拉丁文,原指一種動物來源的毒素。病毒能增殖、遺傳和演化,因而具有生命最基本的特徵,但至今對它還沒有公認的定義。最初用來識別病毒的性狀,如個體微小、一般在光學顯微鏡下不能看到、可通過細菌所不能通過的過濾器、在人工培養基上不能生長、具有致病性等,現仍有實用意義。但從本質上區分病毒和其他生物的特徵是:①含有單一種核酸(DNA或RNA)的基因組和蛋白質外殼,沒有細胞結構;②在感染細胞的同時或稍後釋放其核酸,然後以核酸複製的方式增殖,而不是以二分裂方式增殖;③嚴格的細胞內寄生性。病毒缺乏獨立的代謝能力,只能在活的宿主細胞中,利用細胞的生物合成機器來複製其核酸併合成由其核酸所編碼的蛋白,最後裝配成完整的、有感染性的病毒單位,即病毒粒。病毒粒是病毒從細胞到細胞或從宿主到宿主傳播的主要形式。
病毒一詞的涵義可以是:指那些在化學組成和增殖方式是獨具特點的,只能在宿主細胞內進行複製的微生物或遺傳單位。它的特點是:只含有一種類型的核酸(DNA或RNA)作為遺傳信息的載體;不含有功能性核糖體或其它細胞器;RNA病毒,全部遺傳信息都在RNA上編碼,這種情況在生物學上是獨特的;體積比細菌小得多,僅含有少數幾種酶類;不能在無生命的培養基中增殖,必須依賴宿主細胞的代謝系統複製自身核酸,合成蛋白質並裝配成完整的病毒顆粒,或稱病毒體(完整的病毒顆粒是指成熟的病毒個體)。
由於病毒的結構和組分簡單,有些病毒又易於培養和定量,因此從20世紀40年代後,病毒始終是分子生物學研究的重要材料。
在實踐方面,病毒的研究對防治人類、植物和動物的疾病作出了重要貢獻。如病毒疫苗的發展,利用昆蟲病毒作為殺蟲劑等。1982年將資料齊全而能分類的病毒劃分為7大群:(雙鏈)dsDNA,有包膜;(雙鏈)dsDNA,無包膜;(單鏈)ssDNA,無包膜;(雙鏈)dsRNA,有包膜;(雙鏈)dsRNA,無包膜;(單鏈)ssRNA,有包膜;(單鏈)ssRNA,無包膜。
病毒感染使用抗生素無效,不推薦感染時使用抗生素。干擾素作為處方藥,不得隨意購買和使用,遵照醫囑使用。
特性
 病毒
病毒一、病毒生命形式的兩重性
1.病毒存在的兩重性病毒的生命活動很特殊,對細胞有絕對的依存性。其存在形式有二:一是細胞外形式,一是細胞內形式。存在於細胞外環境時,則不顯複製活性,但保持感染活性,是病毒體或病毒顆粒形式。進入細胞內則解體釋放出核酸分子(DNA或RNA),借細胞內環境的條件以獨特的生命活動體系進行複製,是為核酸分子形式。
⒉病毒的結晶性與非結晶性病毒可提純為結晶體。我們知道結晶體是一個化學概念,是很多無機化合物存在的一種形式,我們可以認為某些病毒有化學結晶型和生命活動型的兩種形式。
3.顆粒形式與基因形式病毒以顆粒形式存在於細胞之外,此時,只具感染性。一旦感染細胞病毒解體而釋放出核酸基因組,然後才能進行複製和增殖,並產生新的子代病毒。有的病毒基因組整合於細胞基因組,隨細胞的繁殖而增殖,此時病毒即以基因形式增殖,而不是以顆粒形式增殖,這是病毒潛伏感染的一種方式。
二、病毒結構和功能的兩重性
1.標準病毒與缺陷病毒在病毒的增殖過程中,由於其基因組因某種微環境因素的影響或轉錄過程的錯誤而發生突變,以致有裝配不全的病毒顆粒產生,稱為缺陷病毒,產生缺陷病毒的原親代病毒,則稱為標準病毒,缺陷病毒顆粒有干擾標準病毒繁殖的作用。
2.假病毒與真病毒一種細胞有兩種病毒同時感染的情況,在增殖過程中,一種病毒可以穿上本身的外殼,這就是真病毒,是這種病毒的應有“面目”;如果一種病毒的核酸被以另一病毒外殼來編碼,則稱為假病毒,此時一種病毒的本來性質,被另一種病毒的性質所掩蓋。
3.雜種病毒和純種病毒兩種病毒混合感染時,除了出現假型病毒外,還有可能出現病毒核酸重組的情況,即一種病毒顆粒之中,可含有兩種病毒的遺傳物質,此可稱為雜種病毒,這是病毒學中一個相當常見的現象。
三、病毒病理學的兩重性
1.病毒的致病性和非致病性關於致病性和非致病性問題,是同宿主細胞相對而言的,在分子水平、細胞水平和機體水平,可能有不同的含義。在細胞水平有細胞病變作用,但在機體水平可能並不顯示臨床症狀,此可稱為亞臨床感染或不顯感染。
2.病毒感染的急性和慢性病毒感染所致的臨床症狀有急、慢之分,有的病毒一般只表現急性感染而很少表現慢性感染;有的則既有急性過程,也有慢性過程。
對病毒的概念可以是:病毒是代謝上無活性,有感染性,而不一定有致病性的因子,他們小於細胞,但大於大多數大分子,他們無例外地在生活細胞內繁殖,他們含有一個蛋白質或脂蛋白外殼和一種核酸,DNA或RNA,甚至只含有核酸而沒有蛋白質,或只有蛋白質而沒有核酸,它們作為大分子似乎太複雜,作為生物體它們的生理和複製方式又千姿百態。Lwoff在“病毒的概念”一文中強調病毒的特殊性時指出,“病毒應該就是病毒,因為它們是病毒”。
分類
結構分類
 病毒
病毒病毒在自然界分布廣泛,可感染細菌、真菌、植物、動物和人,常引起宿主發病。但在許多情況下,病毒也可與宿主共存而不引起明顯的疾病。
Cafeteriaroenbergensis病毒
Cafeteriaroenbergensis病毒的發現模糊了什麼是活的有機體,以及什麼是非生命之間的界限。Cafeteriaroenbergensis,是一種世界上最大、最複雜的海洋病毒,該病毒主要感染那些吃海洋生態系統中非常重要和分布廣泛的浮游生物的掠食者。大多數的病毒都是輕裝旅行的。它們僅僅攜帶了合成新病毒所需的少量基因,並依賴其宿主的機制來完成剩下的工作。2010年10月,加拿大溫哥華市不列顛哥倫比亞大學的MatthiasG.Fischera和同事發現了一種被稱為Cafeteriaroenbergensis飛病毒——它攜帶了令人難以置信的約73萬個脫氧核糖核酸(DNA)鹼基對,其中包括超過500個類似於基因的區域。2010年10月25日,研究人員在美國《國家科學院院刊》(PNAS)網路版上報告了這一研究成果。這也使得這種病毒成為已知最大的海洋病毒,它甚至比一些細菌所具有的DNA還要多。
這種病毒主要感染那些吃海洋生態系統中非常重要和分布廣泛的浮游生物的掠食者。這種病毒的基因組比一些細胞生物的基因組還大。此外,Cafeteriaroenbergensis病毒成為目前(2010年10月)已知的世界最大海洋病毒和第二大病毒,排名僅次於淡水病毒——多噬棘阿米巴模仿病毒,後者擁有120萬個鹼基對。這種病毒可能還是一大組未知但是具有生態重要性的海洋巨型病毒的代表。
研究人員推測,與較小的病毒相比,例如愛滋病病毒(HⅣ)或皰疹病毒,這種病毒——能夠感染Cafeteriaroenbergensis,後者是一種獵食性的單細胞有機體,能夠捕食海洋中的細菌和其他病毒——在其蛋白質的合成過程中扮演了一個更加積極的角色。研究人員指出,這種病毒擁有大量基因,這些基因通常被活細胞用於修復它們的DNA損傷以及合成蛋白質和糖。它還擁有編碼病毒複製需要但是必須從宿主生物那裡獲取的一些蛋白質的基因。
科學家一般不會把病毒劃歸為活的生物體,這是因為病毒無法獨立複製,但是像這樣的巨大病毒——具有它們自己的蛋白質合成機制以及其他通常在活體細胞中才能夠完成的功能——模糊了什麼是活的有機體,以及什麼是非生命之間的界限。唯一已知的較大病毒能夠感染一種淡水變形蟲,且被認為是一個近親。
流感病毒是根據其表面結構來命名的。H代表了血凝素,它的作用是讓病毒能夠結合宿主細胞;N代表了神經氨酸苷酶,其作用是讓已經自我複製的病毒從細胞中釋放出去。甲型流感中H可分為16個亞型,N可分為9個亞型。所有這些亞型的病毒都曾從鳥類體內分離出來過。
醫學分類
一,呼吸道病毒及腸道病毒,麻疹病毒,脊髓灰質炎病毒,柯薩奇病素二,蟲媒病毒及出血熱病毒
流行性乙型腦炎病毒,登革病毒,出血熱病毒,漢坦病毒,伊波拉病毒
三,狂犬病病毒與逆轉錄病毒
狂犬病病毒,人類免疫缺陷病毒,人類嗜T細胞病毒
四,肝炎病毒
A型肝炎病毒,B型肝炎病毒,C型肝炎病毒,丁型肝炎病毒,戊型肝炎病毒
五,皰疹病毒
單純皰疹病毒,水痘-帶狀皰疹病毒,巨細胞病毒,EBV
六,其他病毒
人類乳頭瘤病毒,輪狀病毒,冠狀病毒,風疹病毒
歷史發現
關於病毒所導致的疾病,早在公元前二至三個世紀的印度和中國就有了關於天花的記錄。但直到19世紀末,病毒才開始逐漸得以發現和鑑定。1884年,法國微生物學家查理斯·尚柏朗(CharlesChamberland)發明了一種細菌無法濾過的過濾器(Chamberland氏燭形濾器,其濾孔孔徑小於細菌的大小),他利用這一過濾器就可以將液體中存在的細菌除去。1892年,俄國生物學家伊凡諾夫斯基(DmitryIvanovsky)在研究菸草花葉病時發現,將感染了花葉病的菸草葉的提取液用燭形濾器過濾後,依然能夠感染其他菸草。於是他提出這種感染性物質可能是細菌所分泌的一種毒素,但他並未深入研究下去。當時,人們認為所有的感染性物質都能夠被過濾除去並且能夠在培養基中生長,這也是疾病的細菌理論(germtheory)的一部分。1898年,荷蘭微生物學家馬丁烏斯·貝傑林克(MartinusBeijerinck)重複了Ivanovsky的實驗,並相信這是一種新的感染性物質。他還觀察到這種病原只在分裂細胞中複製,由於他的實驗沒有顯示這種病原的顆粒形態,因此他稱之為contagiumvivumfluidum(可溶的活菌)並進一步命名為virus(病毒)。貝傑林克認為病毒是以液態形式存在的(但這一看法後來被溫德爾·梅雷迪思·斯坦利推翻,他證明了病毒是顆粒狀的)。同樣在1899年,FriedrichLoeffler和PaulFrosch發現患口蹄疫動物淋巴液中含有能通過濾器的感染性物質,由於經過了高度的稀釋,排除了其為毒素的可能性;他們推論這種感染性物質能夠自我複製。20世紀早期,英國細菌學家FrederickTwort發現了可以感染細菌的病毒,並稱之為噬菌體。[14]隨後法裔加拿大微生物學家Félixd'Herelle描述了噬菌體的特性:將其加入長滿細菌的瓊脂固體培養基上,一段時間後會出現由於細菌死亡而留下的空斑。高濃度的病毒懸液會使培養基上的細菌全部死亡,但通過精確的稀釋,可以產生可辨認的空斑。通過計算空斑的數量,再乘以稀釋倍數就可以得出溶液中病毒的個數。他們的工作揭開了現代病毒學研究的序幕。
在19世紀末,病毒的特性被認為是感染性、可濾過性和需要活的宿主,也就意味著病毒只能在動物或植物體內生長。1906年,哈里森發明了在淋巴液中進行組織生長的方法;接著在1913年,E.Steinhardt、C.Israeli和R.A.Lambert利用這一方法在豚鼠角膜組織中成功培養了牛痘苗病毒,突破了病毒需要體內生長的限制。[16]1928年,H.B.Maitland和M.C.Maitland有了更進一步的突破,他們利用切碎的母雞腎臟的懸液對牛痘苗病毒進行了培養。他們的方法在1950年代得以廣泛套用於脊髓灰質炎病毒疫苗的大規模生產。
美國科學家溫德爾·斯坦利1931年,德國工程師恩斯特·魯斯卡和馬克斯·克諾爾發明了電子顯微鏡,使得研究者首次得到了病毒形態的照片。1935年,美國生物化學家和病毒學家溫德爾·梅雷迪思·斯坦利發現菸草花葉病毒大部分是由蛋白質所組成的,並得到病毒晶體。隨後,他將病毒成功地分離為蛋白質部分和RNA部分。溫德爾·斯坦利也因為他的這些發現而獲得了1946年的諾貝爾化學獎。菸草花葉病毒是第一個被結晶的病毒,從而可以通過X射線晶體學的方法來得到其結構細節。第一張病毒的X射線衍射照片是由Bernal和Fankuchen於1941年所拍攝的。1955年,通過分析病毒的衍射照片,羅莎琳·富蘭克林揭示了病毒的整體結構。同年,HeinzFraenkel-Conrat和RobleyWilliams發現將分離純化的菸草花葉病毒RNA和衣殼蛋白混合在一起後,可以重新組裝成具有感染性的病毒,這也揭示了這一簡單的機制很可能就是病毒在它們的宿主細胞內的組裝過程。
20世紀的下半葉是發現病毒的黃金時代,大多數能夠感染動物、植物或細菌的病毒在這數十年間被發現。1957年,馬動脈炎病毒和導致牛病毒性腹瀉的病毒(一種瘟病毒)被發現;1963年,巴魯克·塞繆爾·布隆伯格發現了B型肝炎病毒;1965年,霍華德·馬丁·特明發現並描述了第一種逆轉錄病毒;這類病毒將RNA逆轉錄為DNA的關鍵酶,逆轉錄酶在1970年由霍華德·特明和戴維·巴爾的摩分別獨立鑑定出來。[28]1983年,法國巴斯德研究院的呂克·蒙塔尼和他的同事弗朗索瓦絲·巴爾-西諾西首次分離得到了一種逆轉錄病毒,也就是現在世人皆知的愛滋病毒(HⅣ)。其二人也因此與發現了能夠導致子宮頸癌的人乳頭狀瘤病毒的德國科學家哈拉爾德·楚爾·豪森分享了2008年的諾貝爾生理學與醫學獎。
培養檢測
病毒研究的發展常常與病毒培養和檢測方法的進步有密切的關係,特別在脊椎動物病毒方面,小鼠和雞胚接種、組織培養、超速離心、凝膠電泳、電子顯微鏡和免疫測定等技術,對病毒學的發展具有深刻的影響。噬菌體的培養和檢測方法最為簡單。將噬菌體接種到易感細菌的肉湯培養物中,經18~24小時後,混濁的培養物重新透明,此時細菌被裂解,大量噬菌體被釋放到肉湯中,再經除菌過濾,即為粗製噬菌體。為了測定其中噬菌體的數量,將粗製噬菌體稀釋到每一接種量含100個左右,與過量的細菌混合,然後鋪種於瓊脂平皿上,在溫箱中培養過夜,細菌繁殖成乳白色襯底,被噬菌體裂解的區域則在此襯底上表現為圓形的透明斑,稱為噬斑。噬斑數代表該接種量中有活力的噬菌體數量。如果挑出單個噬斑來培養,就能獲得由單個噬菌體所繁殖的後代,達到分離純化的目的。
動物病毒(見脊椎動物病毒)的培養可在自然宿主、實驗動物、雞胚或細胞培養中進行,以死亡、發病或病變等作為病毒繁殖的直接指標,或以血細胞凝集、抗原測定等作為間接指標。收穫發病動物的組織磨成懸液或有病變的細胞培養液,即為粗製病毒。測定活病毒數量可採用空斑法,其原理與噬斑法相同,但以易感的動物單層細胞代替細菌,在接種適當稀釋的病毒後,用含有培養液和中性紅的瓊脂覆蓋,使病毒感染局限在小面積內形成病變區,襯底的健康細胞被中性紅染成紅色,病變區不染色而顯示為空斑。
至今植物病毒的培養和檢測大都是在整株植物上進行的。從搗碎的病葉汁中製備病毒,常用枯斑法檢測。用手指蘸上混有金剛砂的稀釋病毒在植物葉片上軒輕摩擦,經一定時間後出現單個分開的圓形壞死或退綠斑點,稱為枯斑。
除了利用病毒的致病性定量檢測病毒外,還可套用物理方法,如在電子顯微鏡下計數病毒顆粒,或用紫外分光光度計測定提純病毒的蛋白和核酸量,這些方法所測得的數據包括了有感染性和無感染性的病毒粒。
套用電子顯微鏡不但能看清病毒粒的大小、形態,還可以分辨其表面的蛋白亞單位和內部的核殼等超微結構。DNA
大小與形態
不同病毒的大小變動於20~450納米之間。最大的為痘病毒科,大小為(170~260)×(300~450)納米,最小的為雙聯病毒科,直徑18~20納米。
病毒的形態也是多樣的:球狀(包括二十面體),如脊髓灰質炎病毒和有包膜的如皰疹病毒;桿狀(包括棒狀),如菸草花葉病毒;絲狀,如甜菜黃花病毒;彈狀,如水皰性口炎病毒;複雜構型,如蝌蚪狀的T偶數噬菌體。有些病毒在細胞內呈自然晶體排列。
化學組成
核酸是帶有遺傳密碼的病毒基因組。病毒依所含核酸種類不同可分為DNA病毒和RNA病毒。動物病毒或含DNA,或含RNA;植物病毒除少數組外大多為RNA病毒;噬菌體除少數科外大多為DNA病毒。DNA或RNA可以是線型的或環狀的,可以是單鏈的或雙鏈的。RNA可以分節段或不分節段,單鏈RNA又分正鏈的和負鏈的。
在分節段的RNA植物病毒中,常見多分體基因組,即同一病毒的幾個RNA節段分別裝入衣殼中,形成大小不同的顆粒,有的分裝在兩種顆粒中稱二分體基因組,如豇豆花葉病毒;有的分裝在3種顆粒中稱三分體基因組,如黃瓜花葉病毒和雀麥花葉病毒。
通過遺傳學和生物化學方法,已查明一些病毒的基因圖譜。對MS2和ΦΧ174噬菌體。花椰菜花葉病毒、SV40和B型肝炎病毒核酸的核苷酸序列,已全部查明。
①蛋白質病毒的主要組分,依其功能可分為衣殼蛋白、膜蛋白、糖蛋白和內在酶4類。
衣殼蛋白包裹核酸形成保護性的外殼。簡單的病毒只有1種衣殼蛋白,較複雜的如腺病毒衣殼是由六鄰體、五鄰體和纖維3種蛋白構成的。在有包膜的病毒如流感和水皰性口炎病毒中,膜蛋白一方面與外層脂質相連結,另一方面又同內部的核殼相連結,起到維繫病毒內外結構的作用。糖蛋白位於包膜表面,有的形成突起,如流感病毒的血凝素,能與細胞膜受體結合。病毒雖無完整的酶系統,但常含有一些特殊的酶,如流感病毒的神經氨酸酶和噬菌體的溶菌酶。此外,呼腸孤病毒科、彈狀病毒科、正粘病毒科和副粘病毒科病毒粒中含RNA多聚酶,反錄病毒科含反轉錄酶,均與核酸複製有關。已查明十幾種病毒蛋白的全胺基酸序列。
②脂質存在於包膜中,包膜是在病毒成熟時從細胞質膜或核膜芽生獲得的,所以病毒脂質常具有宿主細胞脂質的特徵。用有機溶劑或去污劑破壞包膜脂質,可使病毒粒裂解。
③糖除核酸中的戊糖外,病毒包膜還含有與蛋白或脂質結合的多糖。
菸草花葉病毒、流感病毒和枯草桿菌噬菌體的電子顯微鏡照片和結構模式圖(見植物病毒、正粘病毒科和細菌病毒)。
複製
病毒複製指病毒粒入侵宿主細胞到最後細胞釋放子代毒粒的全過程,包括吸附、進入與脫殼、病毒早期基因表達、核酸複製、晚期基因表達、裝配和釋放等步驟。各步的細節因病毒而異。
吸附與進入
T4噬菌體先以其尾絲與大腸桿菌表面受體結合,隨後尾鞘收縮,裸露出的尾軸穿入細菌外壁,把頭部內儲存的DNA注射到細菌體內。動物病毒也是先與細胞受體結合,以後或是靠細胞的吞噬作用進入,或是病毒包膜與細胞質膜融合後使核殼進入。植物病毒則是通過傷口侵入或通過媒介昆蟲直接注入。一般情況下,病毒均須經脫殼,即脫去外被的蛋白質釋放核酸,才能進行下一步複製。
基因表達
將其核酸上的遺傳信息轉錄成信使核糖核酸(mRNA),然後再翻譯成蛋白質。一般在核酸複製以前的稱早期基因表達,所產生的早期蛋白質,有的是核酸複製所需的酶,有的能抑制細胞核酸和蛋白質的合成;在核酸複製開始以後的稱晚期基因表達,所產生的晚期蛋白質主要是構成毒粒的結構蛋白質。早期和晚期蛋白質中都包括一些對病毒複製起調控作用的蛋白質。
轉錄
因病毒核酸的類型而異,共有6種方式:雙鏈DNA(dsDNA)的病毒如SV40,其轉錄方式與宿主細胞相同;含單鏈DNA(ssDNA)的病毒如小DNA病毒科,需要通過雙鏈階段後再轉錄出mRNA;含單鏈正鏈RNA(ss+RNA)的病毒如脊髓灰質炎病毒、菸草花葉病毒和Qβ噬菌體,其RNA可直接作為信使,利用宿主的蛋白質合成機器合成它所編碼的蛋白質;含單鏈負鏈RNA(ss-RNA)的病毒如水皰性口炎病毒和流感病毒,需先轉錄成互補的正鏈作為其mRNA,ssRNA的反錄病毒如雞肉瘤病毒和白血病病毒,需先經反轉錄成dsDNA而整含到宿主染色體中,於表達時再轉錄成mRNA,含dsRNA的呼腸孤病毒,則以保守型複製方式轉錄出與原來雙鏈中的正鏈相同的mRNA。
有些病毒(如腺病毒和SV40)的基因是不連續的,有外顯子與內含子之分,轉錄後有剪接過程,把內含子剪除而把外顯子連線起來,才有mRNA的功能。多數病毒的mRNA還需經過其他加工,如在5′端加上“帽子”結構和在3′端加上多聚腺嘌呤核苷酸。
病毒基因轉錄所需酶的來源也不相同,如小DNA病毒科、乳多泡病毒科所需依賴於DNA的RNA多聚酶,都是利用宿主原有的酶;而彈狀病毒科、正粘病毒科、副粘病毒科和呼腸孤病毒科所需的依賴於RNA的RNA多聚酶,以及反錄病毒科所需的反轉錄酶,都是病毒粒自備的。
翻譯
不同病毒mRNA翻譯的方式是不同的。一般認為噬菌體的翻譯是多順反子的,如Qβ的RNA上有3個順反子(為單個肽鏈編碼的基因功能單位),可沿著1條mRNA獨立地翻譯出3種多肽。動物病毒的翻譯是單順反子的,即由其基因組轉錄成不同的mRNA,每種mRNA翻譯成一種多肽。分節段基因組病毒如流感病毒和呼腸孤病毒,每1節段RNA構成1個順反子,多分體基因組的植物病毒也是如此。脊髓灰質炎病毒的mRNA先被翻譯成1個分子量為20萬的巨肽,再經裂解成為衣殼蛋白和酶。
有些病毒如ΦΧ174,Qβ噬菌體和SV40等,存在基因重疊現象,即按讀碼位相不同而從同一核苷酸序列可以表達出一種以上的蛋白質。這是病毒經濟地利用其有限的遺傳信息的1種方式。
核酸複製
DNA病毒按照經典的沃森-克里克鹼基配對方式進行DNA複製。乳多泡病毒的環狀DNA按“滾環”模式進行複製時,需要有核酸內切酶和連線酶參與。病毒RNA是通過半保留方式複製的,即以病毒RNA(vRNA)為模板,同時轉錄幾個互補鏈(cRNA),cRNA轉錄完成並脫落後,又以同樣方式再轉錄出新的vRNA。因此,在感染細胞中可以查出具有部分雙鏈結構而又拖著多條長短不同單鏈“尾巴”(正在合成中的互補鏈)的“複製中間體”。
病毒核酸複製所需酶的來源也各不相同。SV40DNA合成所需的酶都來自宿主。含RNA的Qβ噬菌體、小RNA病毒科和含ssRNA的植物病毒所需RNA多聚酶的某個亞基,可能由病毒基因編碼,而其他亞基來自宿主。皰疹病毒DNA複製所需的酶,部分地由病毒編碼,如DNA多聚酶和胸苷激酶,可能還有核苷酸還原酶。痘類病毒的獨立自主能力最強,甚至能在去核細胞中進行DNA複製,其基因組至少能為75種蛋白質編碼,包括DNA多聚酶、胸苷激酶、脫氧核糖核酸酶和聚核苷酸連線酶。
裝配與釋放
病毒核酸和結構蛋白是分別複製的,然後裝配成完整的病毒粒。最簡單的裝配方式(如菸草花葉病毒)是核酸與衣殼蛋白相互識別,由衣殼亞單位按一定方式圍繞RNA聚集而成,不藉助酶,也無需能量再生體系。許多二十面體病毒粒先聚集其衣殼,然後再裝入核酸。有包膜的病毒,在細胞內形成核完後轉移至被病毒修飾了的細胞核膜或質膜下面,以芽生方式釋放病毒粒。T4噬菌體則先分別裝配頭部、尾部和尾絲,最後組合成完整病毒粒,裂解細菌而釋放,其中有些步驟需酶的作用。
細胞水平上的感染類型和宿主反應
很早發現噬菌體感染有裂解性和溶源性之分。以大腸桿菌的λ噬菌體為例,裂解性感染於經歷上述複製周期後產生大量子代病毒粒而將細菌裂解;而溶源性感染時,噬菌體DNA環化並整合到大腸桿菌DNA的特異性位點上,隨著細菌的分裂而傳給子代細菌,細菌不被裂解也不產生子代病毒粒。營養條件、紫外線或化學藥物都能使溶性源感染轉化為裂解性。動物的DNA病毒如SV40、腺病毒、皰疹病毒等於感染敏感細胞(稱為容許細胞)後,形成裂解性感染,而於感染不大敏感的細胞(稱為不容許細胞)後,則形成轉化性感染。轉化性感染與溶源性感染相似,病毒DNA或其片段整合於細胞染色體上,並隨細胞分裂而傳給子代細胞,表達其部分基因(一般為早期基因),但不產生子代病毒粒,細胞也不死亡,但被轉化成類似於腫瘤細胞,可無限地傳代。另一方面,RNA腫瘤病毒(如雞肉瘤病毒)必須先將其RNA反轉錄成dsDNA並整合到細胞染色體上,才能進行複製,所以這種感染方式是獨特的,既是轉化性感染,又產生大量病毒粒。
宿主細胞對病毒感染的反應有4種:無明顯反應、細胞死亡、細胞增生後死亡和細胞轉化。例如,副粘病毒SV5在細胞培養中產生大量病毒而不引起明顯反應。多數病毒感染敏感細胞時,由於抑制了細胞核酸和蛋白質合成而引起細胞死亡。痘病毒感染時,先刺激細胞多次分裂然後死亡,造成痘皰病灶。DNA病毒和RNA腫瘤病毒則引起細胞轉化。
有些動物病毒於感染宿主細胞後,在胞核或細胞質內形成具有特殊染色特性的內含物,稱為包涵體,如痘病毒的細胞質內包涵體和皰疹病毒的胞核內包涵體。這些包涵體有的是由未成熟或成熟的病毒粒構成,有的是宿主細胞的反應產物,有的是兩者的混合物。有些昆蟲病毒的病毒粒包埋在蛋白基質中,形成包涵體如核型多角體病毒。
脊椎動物細胞感染病毒後的另一種反應是產生干擾素。干擾素是一種動物細胞編碼的蛋白,其基因平常處於不活動狀態,於病毒感染或經雙鏈RNA誘導後活化。干擾素有廣譜的抗病毒作用,但並不直接作用於病毒,其作用機制是通過與細胞膜結合,激活具有抗病毒作用的3種酶,阻斷了病毒mRNA的翻譯。干擾素在防止病毒擴散和疾病恢復中有一定作用,並有可能成為一種抗病毒藥物。
機體水平上的感染類型和宿主反應
高等動、植物感染病毒後,可表現為顯性感染和持續感染,動物病毒還可表現為隱性感染。隱性感染無臨床症狀,顯性感染表現為臨床疾病;在持續感染中,病毒在機體內長期存在。動物病毒的持續感染又分為潛伏感染、慢性感染和長程感染3類。潛伏感染如皰疹,平常無症狀也查不到病毒,但由於內外因素的刺激而復發時出現病毒;慢性感染如B型肝炎,有或無症狀,但可查到病毒;長程感染限於少數病毒,如綿羊的Maedi-visna(一種反錄病毒感染)可查到病毒;潛伏期和病程都很長,進行性發病直至死亡。
高等動物能對病毒感染產生特異性免疫反應。免疫反應分為體液免疫和細胞免疫兩類,體液免疫表現為由B細胞產生的抗體,其中包括能特異地滅活病毒的中和抗體。中和抗體在預防再感染中起主導作用。細胞免疫的主要表現是識別病毒抗原並發生反應的T淋巴細胞,在清除病毒和病毒感染細胞中起主導作用。
植物細胞對病毒常有過敏反應,細胞迅速死亡,形成枯斑,同時病毒複製也受到限制。另一種反應是產生一種很象干擾素的抗病毒因子,能保護未受感染的細胞。
比如:EV71腸道病毒
據專家介紹,腸道病毒EV71是人腸道病毒的一種,簡稱EV71,一年四季都可發生,常見於4~9月,主要通過唾液、皰疹液、糞便污染的手、毛巾、手絹、牙杯、玩具、食具、奶具及床上用品、內衣等密切接觸傳播,常引起兒童手足口病、病毒性咽峽炎,重症患兒可出現肺水腫、腦炎等,統稱為腸道病毒EV71感染疾病。該病多發生於兒童,尤其3歲以下嬰幼兒多發,少數病情較重,嚴重的會引起死亡。
患兒感染腸道病毒EV71後,多以發熱起病,一般為38℃左右,發熱同時在口腔、手足、臀部出現皮疹,或出現口腔黏膜皰疹。
部分病人早期有咳嗽等感冒樣表現。發熱1~2天后開始出現皮疹,通常出現在手掌和足底,也可以出現在臀部。有的患兒不發熱,只表現為手、足、臀部皮疹或皰疹性咽峽炎,病情較輕。大多數患兒在一周以內體溫下降、皮疹消退,病情恢復。
中國疾病預防控制中心專家介紹說,如果發現孩子發燒、有皮疹等症狀,儘快到正規醫院就診。孩子患病後應暫停去幼稚園和學校,避免傳染給他人,防止再感染其他疾病。
專家強調,患兒的家庭應使用肥皂、84消毒液對日常用品、玩具、尿布進行消毒,對奶具、餐具煮沸消毒。患兒糞便及其他排泄物可用消毒劑或漂白粉消毒;將衣被陽光暴曬,室內保持通風換氣。
春夏是腸道病毒感染容易發生的季節,要講究環境衛生、食品衛生和個人衛生。不喝生水、不吃生冷食物,飯前便後洗手,保持室內空氣流通。儘量不要帶嬰幼兒去人群密集的場所。哺乳的母親要勤洗澡、勤換衣服,餵奶前要清洗奶頭。
在托幼機構、國小等兒童集體生活、學習的場所,專家建議要做好晨間體檢,發現有發熱、皮疹的孩子,要立即要求家長帶小孩去醫院就診,同時報告相關部門。如發現有發熱、皮疹的孩子後,要立即對玩具、被褥、桌椅等進行消毒;同時做好食堂、衛生間、教室等的消毒處理。
危害
致瘤作用有一些病毒能誘發良性腫瘤,如痘病毒科的兔纖維瘤病毒、人傳染性軟疣病毒和乳多泡病毒科的乳頭瘤病毒;另有一些能誘發惡性腫瘤,按其核酸種類可分為DNA腫瘤病毒和RNA腫瘤病毒。DNA腫瘤病毒包括乳多泡病毒料的SV40和多瘤病毒,以及腺病毒科和皰疹病毒科的某些成員,從腫瘤細胞中可查出病毒核酸或其片段和病毒編碼的蛋白,但一般沒有完整的病毒粒。RNA腫瘤病毒均屬反錄病毒科,包括雞和小鼠的白血病和肉瘤病毒,從腫瘤細胞中可查到病毒粒。這兩類病毒均能在體外轉化細胞。在人類腫瘤中,已證明EB病毒與伯基特淋巴瘤和鼻咽癌有密切關係;從一種T細胞白血病查到反錄病毒。此外,Ⅱ型皰疹病毒可能與宮頸癌病因有關,B型肝炎病毒可能與肝癌病因有關。但是,病毒大概不是唯一的病因,環境和遺傳因素可能起協同作用。
病毒感染常發生在感冒等上呼吸道感染後,病毒顆粒可由血循環直接進入內耳血循環中,引起耳蝸毛細胞、神經節細胞及微血管等結構的破壞。病毒亦可經圓窗侵入內耳,引起迷路炎等病損,引起耳聾。
起源
對於病毒的起源曾有過種種推測;一種觀點認為病毒可能類似於最原始的生命;另一種認為病毒可能是從細菌退化而來,由於寄生性的高度發展而逐步喪失了獨立生活的能力,例如由腐生菌→寄生菌→細胞內寄生菌→支原體→立克次氏體→衣原體→大病毒→小病毒;還有一種則認為病毒可能是宿主細胞的產物。這些推測各有一定的依據,因此病毒在生物進化中的地位是未定的。但是,不論其原始起源如何,病毒一旦產生以後,同其他生物一樣,能通過變異和自然選擇而演化。
分類
病毒分類命名的工作現由國際病毒分類委員會負責,已於1971.1976.1979和1982年發表過4次報告。
1982年將資料較齊全而能分類的病毒劃分為7大群,分群的根據是基因組的核酸種類(DNA或RNA)、類型(ds或ss)和有無包膜。
7大群中包括59個科組:
dsDNA,有包膜4科
dsDNA,無包膜8科,1組
ssDNA,無包膜3科,1組
dsRNA,有包膜1科
dsRNA,無包膜1科,4個可能科
ssRNA,有包膜8科,1組
ssRNA,無包膜4科,22組,1個可能組
如按宿主分類,則為:
細菌病毒10科
真菌病毒3個可能科
植物病毒24組,1個可能組
無脊椎動物病毒2科,1組
脊椎動物病毒9科
無脊椎、脊椎動物共有的病毒有6科,即痘病毒科虹彩病毒科、小DNA病毒科、披膜病毒科、布尼亞病毒科和小RNA病毒科,以及一個可能科,即二節段雙鏈RNA病毒。
無脊椎、脊椎動物和植物共有的病毒有2科,即呼腸孤病毒科和彈狀病毒科。
病毒分類還處於初期階段,以後還會迅速發展和演變。對資料較齊全的動物病毒和噬菌體都已立為科,科名採用拉丁文;而植物病毒則只立組,組名多採用縮拼法,即將某科的典型代表病毒的普通名稱如Tobaccomo-saicvirus縮拼為Tobamo-virus。科下分亞科及屬,屬下即為各個病毒的普通名稱,目前尚未分種。
作用價值
生物病毒的好處1.噬菌體可以作為防治某些疾病的特效藥,例如燒傷病人在患處塗抹綠濃桿菌噬菌體稀釋液
2.在細胞工程中,某些病毒可以作為細胞融合的助融劑,例如仙台病毒
3.在基因工程中,病毒可以作為目的基因的載體,使之被拼接在目標細胞的染色體上
4.在專一的細菌培養基中添加的病毒可以除雜
5.病毒可以作為精確制導藥物的載體
6.病毒可以作為特效殺蟲劑
7.病毒還在生物圈的物質循環和能量交流中起到關鍵作用.
8.病毒還可以用來治療疾病,比如癌症
病毒疫苗對人類有防病毒有好處--促進了人類的進化,人類的很多基因都是從病毒中得到的。
病毒是一種非細胞生命形態,它由一個核酸長鏈和蛋白質外殼構成,病毒沒有自己的代謝機構,沒有酶系統。因此病毒離開了宿主細胞,就成了沒有任何生命活動、也不能獨立自我繁殖的化學物質。一旦進入宿主細胞後,它就可以利用細胞中的物質和能量以及複製、轉錄和轉譯的能力,按照它自己的核酸所包含的遺傳信息產生和它一樣的新一代病毒。
病毒基因同其他生物的基因一樣,也可以發生突變和重組,因此也是可以演化的。因為病毒沒有獨立的代謝機構,不能獨立的繁殖,因此被認為是一種不完整的生命形態。科學家發現了比病毒還要簡單的類病毒,它是小的RNA分子,沒有蛋白質外殼,但它可以在動物身上造成疾病。這些不完整的生命形態的存在說明無生命與有生命之間沒有不可逾越的鴻溝。
不同尋常的自然角色:
其實,病毒也並非一無是處,它在人類生存和進化的過程當中,扮演了不同尋常的角色,人和脊椎動物直接從病毒那裡獲得了100多種基因,而且人類自身複製DNA的酶系統,也可能來自於病毒。
生活場所
嚴格的活細胞內寄生
新型病毒
專家稱SARS病毒不是“非典”總部位於瑞士日內瓦的世界衛生組織於當地時間24日發布公告稱,一名感染了類似“非典”(Atypicalpneumonias,簡稱ATP)病毒的卡達男子目前正在英國接受特別治療。
此前,已有一名沙特籍男子因感染相同病毒而死亡。世衛組織尚未針對新病毒發布旅行警告,稱正進行進一步調查。
病毒與癌
第一個發現癌的人弗朗西斯·佩頓·勞斯弗朗西斯·佩頓·勞斯(FrancisPeytonRous)1879年10月5日出生於美國,是紐約市洛克菲勒研究所的內科醫生和病毒學家。
勞斯醫生畢業於馬里蘭州巴爾的摩市約翰斯·霍普金斯大學。1911年1月21日,弗朗西斯·佩頓·勞斯發表了一份報告:癌性腫瘤是病毒所致。這一提法在醫學史上是首次。因為還沒有證據表明癌症對人或動物有傳染性。勞斯也成為發現這種“腫瘤病毒”的第一人,因為這種病毒最先是在那隻被勞斯接診的雞身上發現的,所以病毒被命名為“勞斯雞肉瘤病毒”。1966年,已經87歲高齡的勞斯在距離發現這種病毒55年之後,獲得了諾貝爾生理學或醫學獎。這種病毒的發現與勞斯積極的工作是分不開的,多年來,勞斯一直在積極地進行著研究工作,事實上,直到他過90歲生日時為止,他一直都在工作。
愛滋進展
功能性治癒愛滋嬰兒病毒美國研究人員3日報告說,他們通過抗逆轉錄病毒療法實現了首次“功能性治癒”愛滋病病毒嬰兒感染者。
美國約翰斯·霍普金斯兒童醫療中心、密西西比大學等機構的研究人員3日在美國亞特蘭大舉行的“2013年逆轉錄病毒與機會性感染大會”上報告說,他們在兩年前選取一名通過母嬰傳播感染愛滋病病毒的女嬰為治療對象。在該嬰兒出生30小時後,研究人員對其進行組合式抗逆轉錄病毒治療。檢測證明,經過治療該女嬰血液中愛滋病病毒的數量明顯遞減,在其出生29天后,體內的愛滋病病毒已經檢測不到。研究人員在隨後18個月內繼續對她進行抗逆轉錄治療,並在停止治療10個月後發現,其體內的愛滋病病毒抗體仍為陰性,常規血液檢測中未發現愛滋病病毒存在。
研究人員說:“對新生兒進行抗逆轉錄病毒治療後可以阻止體內藏匿的愛滋病病毒感染宿主細胞,該療法能夠清除、抑制該病毒,在非終身治療的情況下實現‘功能性治癒’。”
“功能性治癒”是指感染者體內的愛滋病病毒被完全抑制,機體免疫功能正常,即便不接受治療,用常規方法也難以在患兒血液中檢測出病毒。
研究人員表示,根除愛滋病病毒,即“根本性治癒”愛滋病當前難以實現。針對愛滋病病毒嬰兒感染者的藥物治療,一般開始於其出生後3到4個月之間,因此儘早、準確進行抗逆轉錄治療對感染愛滋病病毒的嬰兒意義重大。
