
名詞起源
 人類的進化
人類的進化達爾文並未對“evolution”下過定義,在1859年出版的《物種起源》第一版中,也並未使用“evolution”這個字。當時達爾文是使用“經過改變的繼承”(descentwithmodification)、“改變過程”(processofmodification)或是“物種改變的原理”(doctrineofthemodificationofspecies)等。evolution這個字在當時生物學上的意義,指的是胚胎髮育的過程,並且在當時的一般用語中具有“進步”的意含,而達爾文反對將“進步”之類的用語來描述生物改變的過程。他曾在《物種起源》第7章中說:“(天擇)的最後結果,包括了生物體的進步(advance)及退步(retrogression)兩種現象”。而後來包括達爾文在內,之所以改用evolution來描述生物演化現象,是經由英國哲學家赫伯特•史賓賽在許多著作里進行的名詞統一。
進化與演化
原本中文對“evolution”這個字有兩種翻譯。“進化”一詞是來自日語(日制漢語)。而嚴復則是最早反對使用“進化”的人之一,後人在《天演論》書尾的名詞表中寫到:“evolution一詞,嚴氏譯為天演,近人撰述多以進化二字當之。赫胥黎於本書導言中實嘗有一節,立evolution之界說;謂為初指進化而言,繼則兼包退化之義。嚴氏於此節略而未譯,然其用天演兩字,固守赫氏之說也”。也就是說,嚴復主張以“天演”取代“進化”。
根據台灣教育部所編輯的辭典,“進化”定義為生物由低級到高級、由簡單到複雜的發展過程,並將“退化”定義為進化的反義詞。而“演化”則定義為生物物種為了因應時空的嬗變,而在形態和與行為上與遠祖有所差異的現象。
目前中文對於如何翻譯“evolution”仍有爭議。支持使用“演化”的學者認為,演化在字面上的意義比較中性,能表達連續與隨機的意義;進化則帶有“進步”的含意。而且由於漢語中“進”與“退”是代表相反意義的兩個字,因此若使用進化,則在邏輯上不易將“退化”定義為進化的一種類型。對翻譯的爭論也表現了人們對進化論理解的變化,過去“進化”多表示生物朝適應環境的方向演化,而當前多人為生物的演化是隨機的,並沒有進步退步之分。
演化模型
微觀演化與巨觀演化
演化可以依據時間長短與差異程度,分成“微觀演化”(微演化)與“巨觀演化”(廣演化)。微觀演化指幾個世代中,基因頻率小範圍的變化,例如現今世界各地人類的差異。巨觀演化指長時間的演化過程,例如人類與滅絕祖先的關係。巨觀演化的歷史中可能包括生物群在化石紀錄中的突然出現、丟失的環節、物種長期停滯等難以解釋的現象。例如寒武紀地層中保存大量化石的現象。
演化速率
關於巨觀演化的過程,有許多不同的理論,主要是用來解釋化石紀錄中難以解釋的現象。傳統的觀念認為,許多微觀演化的累積,經歷足夠時間之後便形成新物種,這樣的理論稱為漸變論。而漸變論又有許多形式,例如由史蒂芬•古爾德與尼爾斯•艾崔奇在1972年提出的疾變平衡論。這種理論認為生物的演化歷史是許多走走停停的過程,在大多數時間並沒有太大的變化。而某些個體中存在關鍵基因(如同源異形盒),可能導致新物種迅速形成並大量繁殖,之後再恢復平衡。如此便能夠合理的解釋化石紀錄不平均的問題。其實古爾德經常以批判漸變論的姿態出現在大眾媒體,但是道金斯認為,疾變平衡論只是漸變論的一種形式。而極端的漸變論,認為演化過程是以等速進行,對於化石數量的不平均,則以“化石紀錄本身並不完整”來解釋。極端的漸變論,仍然經常使用在分子層次,並發展出分子時鐘的觀念。但是近年的研究發現,分子時鐘也並非等速進行,而是在不同物種或是不同年代具有不同的演化速度。且不同的研究中,對分子演化速率的推算差異相當大。
另外有一種躍進論,認為物種只需要一個世代就能夠形成,且可能具有創造論與災變論等形式。創造論與災變論都是源自宗教,如《創世紀》與大洪水,不過現在的災變論已將火山爆發、隕石撞擊等所有來自地球內外的影響包含在內。
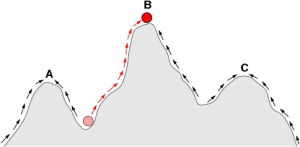 生殖成就空間一般會顯現出山峰與山谷,山峰顯示生物所能達到的最高生殖成就,山谷則顯示其區隔,解釋特徵差異的存在。生殖成就空間除了可以如此圖般以二維方式呈現,也可以三維方式表達。圖中直向為生殖成就,橫向為表現型變數。
生殖成就空間一般會顯現出山峰與山谷,山峰顯示生物所能達到的最高生殖成就,山谷則顯示其區隔,解釋特徵差異的存在。生殖成就空間除了可以如此圖般以二維方式呈現,也可以三維方式表達。圖中直向為生殖成就,橫向為表現型變數。數學模式
生殖成就空間一般會顯現出山峰與山谷,山峰顯示生物所能達到的最高生殖成就,山谷則顯示其區隔,解釋特徵差異的存在。生殖成就空間除了可以如此圖般以二維方式呈現,也可以三維方式表達。圖中直向為生殖成就,橫向為表現型變數。
生殖成就空間一般會顯現出山峰與山谷,山峰顯示生物所能達到的最高生殖成就,山谷則顯示其區隔,解釋特徵差異的存在。生殖成就空間除了可以如此圖般以二維方式呈現,也可以三維方式表達。圖中直向為生殖成就,橫向為表現型變數。
生物的變異可以量化為數字,因此也能夠建立出數學模型。較早的數值分類學(numericaltaxonomy)將生物的特徵量化為數字,並且依照這些數字進行分類,藉此找出它們的親緣關係。
現在的科學家一般認為生物型體的演化,是源自於基因的變異(基因中心演化觀點)。而且由於基因突變具有一些規律性,因此複雜的演化過程,可以簡化成數學模型。早期的新達爾文主義科學家使用線性的模型,例如將為每一個對偶基因定下一個“天擇係數”,藉此估計此對偶基因在每一世代中的基因頻率。現今則多用非線性的方式分析,例如一種稱為生殖成就空間(fitnesslandscape)的曲面圖形,可以用來表達生物個體在繁殖上的能力,與其特徵之間的相關性。由圖形可以看出具有不同特徵的生物可能同時皆有較高的生殖成就。
由於這些生物的變異可以化成許多種不同的變數,因此數學上用來處理多維空間的技巧,便可套用在生物關係的分析。此外這些數學方法的優點是計算過程上相當精確,缺點是其正確性取決於人為的判斷與假設。
進化證據
古生物學與化石紀錄
 三葉蟲化石,三葉蟲是蝦與蟹的親戚,活躍於大約5億年前到2億五千萬年前
三葉蟲化石,三葉蟲是蝦與蟹的親戚,活躍於大約5億年前到2億五千萬年前古生物學是以生物化石為基礎,以研究生物親緣關係的一種研究。當生物個體死亡之後,它的屍體通常會經由微生物分解而腐化,使得生存痕跡消失。但有時候這些遺蹟可能會因為某些因素而被保存。只要是來自古代生物造成的痕跡,或是生物體本身,都可以稱為化石。化石對於了解生物演化歷程而言相當重要,因為化石是較為直接的證據,且帶有許多詳細的資訊。
在化石形成過程中,生物體外的痕跡由於快速地受到掩埋,因此不會發生風化與分解的情形。而較為常見的化石,則通常源自骨骼或外殼等堅硬部位,並經由類似鑄模的過程形成。堅硬的骨骼在動物死亡之後,會因為有機物的腐敗,而產生一些漏洞。將骨骼掩埋的砂石或礦物,則會經由這些漏洞侵入骨骼內部,並將其填滿。這種過程稱為置換作用,屬於型體的保留,而不是生物體本身的保留。也有一些化石是生物體本身,例如被冰凍的猛獁象、琥珀里的昆蟲。此外,古代動物的腳印,或是植物在地底下因為溫度與壓力的作用而碳化,都可稱為化石。
不同時代的生物化石,會出現的在不同的地層中,如此便能夠研究古生物之間,以及它們與現代生物之間的關係。“失落的環結”指演化過程可能出現過,卻尚未發現的物種;而連線兩個物種之間的化石,則稱為“過渡化石”。例如可能位在鳥類與恐龍中間的始祖鳥(Archaeopteryx)化石;以及最近所發現的一種具有四肢的大型淺水魚(Tiktaalik),可能是魚類與兩棲類的過渡化石。
化石紀錄對於古生物的研究有所限制,因為形成化石並不容易。舉例而言,軟體動物身上並沒有太多能夠形成化石的部分,還有一些生物生存在難以形成化石的環境當中。即使化石形成之後,也有可能因為某些原因被摧毀,使得大多數化石皆是零散的狀態,只有少數化石能夠保持完整。而當演化上的改變在族群當中只占有少部分,或是環境變化使族群規模縮小,都會使它們形成化石的機率相對較小。此外,化石几乎無法用來研究生物內部器官構造和機制。
生物地理學與物種分布
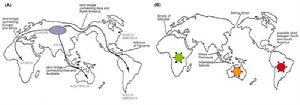 A圖顯示古代大陸間有陸橋連結,因此動物可在大陸間自由遷徙。B圖顯示陸橋消失之後,各地物種受到孤立,並形成新起源的情形。
A圖顯示古代大陸間有陸橋連結,因此動物可在大陸間自由遷徙。B圖顯示陸橋消失之後,各地物種受到孤立,並形成新起源的情形。由於板塊移動造成的大陸漂移(如南美洲與非洲),以及冰河時期前後造成的海平面高度的變化(如白令海峽陸橋),改變了陸地之間的相連性,使得一些相差遙遠的地區,雖然能夠在地底下挖出許多相似的生物化石,如今卻因為海洋或山脈的隔離,而使現有的物種具有相當大的差異。
例如在南美洲的新世界猴、美洲豹與駱馬;以及非洲的舊世界猴、獅子與長頸鹿。此外,與世界上其它地方的胎盤動物相比較為原始的有袋類動物,雖已大多滅絕,但澳大利亞大陸卻依然保留如袋鼠與無尾熊等許多有袋類。除了不同陸塊之間具有這種現象之外,在大陸與其鄰近島嶼,也因為曾經在地理上相連,而能夠找到相似但具有變異的物種。例如台灣、中國大陸與日本的獼猴之間的差異。
型態比較
脊椎動物五趾肢的比較,支持了脊椎動物具有共同祖先的理論。舉例而言,雖然人類、貓、鯨魚與蝙蝠的五趾肢在型態上有所差異,但是主要架構都很相似。這些“同源”的構造,適應了不同的功能,如抓握、行走、游泳與飛行。
此外有一些構造在功能上相似,但卻具有不同的型態。例如蝙蝠、鳥類與昆蟲的翅膀;昆蟲與脊椎動物的腿;章魚與脊椎動物的眼睛;魚類、鯨魚與龍蝦的鰭等。這類“異源”的構造,適應了相同的功能,如飛行、行走、感光與游泳。
發育過程
在發育初期都非常相似,在發育的過程中,這樣的相似會逐漸減少,最後形成各物種的型態。舉例而言,雖然各種成熟的脊椎動物差異很大,但是它們的胚胎型態在發育初期卻非常相似,腮裂仍然出現在已經沒有腮的爬蟲類、鳥類與哺乳類胚胎中。魚類的二房心會被保留到成體,而人類的胚胎雖然也有這種構造,卻會在胚胎成熟後消失。
1870年代提出胚胎重演學說的恩斯特•海克爾,認為不同物種在同一段發育時期的差異也會顯現出這些物種在演化上的親近程度,而且這些生物演化歷史的重複表現,能夠出現在任何生物的胚胎髮育過程。但是到了1997年,海克爾的理論正式被推翻。理察森與先前的一些研究發現,胚胎髮育的型態,並不完全符合演化上的親近程度。即使如此,現今的某些中學生物教科書依然繼續使用海克爾的理論。
在胚胎重演論提出直到被推翻的期間,胚胎學對於演化機制的解釋並沒有太大的進展。但是近年的演化發育生物學(Evolitionarydevelopmentalbiology、EvoDevo)研究,將分子生物學與發育生物學等學門結合,解釋基因的改變對於動物形態的控制過程。同時也發現外表差異相當大的動物之間,也擁有相同的調控基因。以及相同的基因在不同的時間與空間,具有不同的作用。這些調控動物發育過程的基因,主要為一類Hox基因
分子證據
分子生物學與生物化學可以利用構成生命的各種分子,來研究生物的親緣關係。這些分子包括了核糖核酸(RNA)、脫氧核糖核酸(DNA)、蛋白質與糖體等。現在的科學家已經可以用機器與電腦將這些分子的鹼基序列、胺基酸序列定序。研究這些序列的學門分別為基因體學、蛋白質體學。此外還有一種更為複雜的糖體學,專門分析糖類構造。
基因體學透過脫氧核糖核酸序列的比較,可以建立出與傳統分類學大致符合的種系發生樹。這種方法通常用來確認並增強分類學的研究,將其它演化證據的漏洞修補或是修正。例如人類的DNA序列與黑猩猩的差異約為1.2%,與大猩猩的差異約為1.6%,與狒狒則為6.6%,如此就能推斷並量化人類與猿類的親緣關係。此外一種組成核糖體的16SrRNA序列,也是研究親緣關係的重要分子,這些研究導致了三域系統的出現,將真核生物、古生菌與細菌分開。
蛋白質體學也支持了生物具有共同祖先的說法,因為許多生命所需的蛋白質,例如核糖體、DNA聚合酶與RNA聚合酶,不但出現在較原始的細菌,也出在複雜的哺乳類體內。這些蛋白質的核心部分在不同生物中保有相似的構造與功能,而較複雜的生物具有較多的蛋白質次單位(proteinsubunit),以調控更複雜的蛋白質相互作用。
生物體內有一種脫氧核糖核酸片段,雖然與已知基因具有相似的序列,但是卻無法發揮作用並完成蛋白質的製造。這類無用的核酸片段稱為偽基因,是支持生物具有共同祖先的證據之一。它們之所以無法正常作用,是因為在演化過程中,基因突變累積過多的緣故。
抗藥性
微生物與病毒的抗藥性,在醫學上是重要問題,也是演化證據之一。例如金黃葡萄球菌在1943年時仍可使用青黴素(盤尼西林)治療,到了1947年就已經發現具抗藥性的菌株。1960年代改用甲氧苯青黴素,同樣因為抗藥性菌種的散布,使得1980年代改用萬古黴素,2002年時,已發現抗萬古黴素的菌種。
遺傳
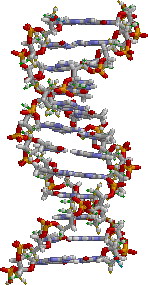 DNA結構,鹼基位於中心,外側環繞著由磷酸根與糖類所形成的雙螺鏇。
DNA結構,鹼基位於中心,外側環繞著由磷酸根與糖類所形成的雙螺鏇。生物體的遺傳是發生在一些不連續性狀上,也就是生物的特定特徵。以人類為例,眼睛的色彩是一個是一項特徵,可遺傳自父母的其中一個。遺傳性狀是由基因所控制,而在生物個體基因組中完整的一套基因,則成為基因型。
完整的一套可觀察性狀,可形成生物的構造或是行為,稱為表現型。這些性狀來自基因型與環境的互動作用。因此生物體的表現型並非完全來自遺傳,例如皮膚的曬黑情況,是決定於個人的基因型與陽光的照射。每個人之所以對陽光有不同的反應,是因為基因型的差異,較顯著的例子是擁有白化性狀的個體,這類個體不會曬黑,且相當容易曬傷。
基因是DNA分子中一些含有遺傳訊息的區域,DNA則是含有四種鹼基的長鏈分子。不同的基因具有不同的鹼基序列,這些序列以編碼形式形成遺傳訊息。細胞里的DNA長鏈會與蛋白質聚集形成一種生為染色體的構造,染色體上的特定位置,則稱作基因座(locus)。有時基因座上的序列在不同個體之間有所差異,這些各式各樣變化型態稱為等位基因(allele)。突變可使基因序列改變,產生新的等位基因。當突變發生虞基因時,新形成的等位基因可能會影響此基因所控制的性狀,使表現型改變。不過單一等位基因對應單一性狀的情形較少,多數的性狀更為複雜,而且是由許多進行互動作用的基因來控制。
進化變異
突變
突變是指遺傳的物質發生改變,廣義的突變包括染色體數目和結構變異。不過一般所說的突變,是核酸序列的改變,也就是基因突變。基因突變是產生遺傳變異的最根本原因,細胞中的遺傳物質(通常是脫氧核糖核酸或核糖核酸)能夠經由許多方式改變,例如細胞分裂時的複製錯誤、放射線的照射、化學物質的影響或是病毒感染。多細胞生物的基因突變,可依照發生的細胞種類分為兩種。生殖細胞突變能夠遺傳到下一代;體細胞突變則通常限制在個體中。
基因突變可能對個體有害,也可能對個體有益,或是兩者兼具。有害的隱性基因因為不會出現症狀而被保留,當這些隱性基因配成對時,就可能使個體得到病變或是死亡。有一些基因雖然可能會造成病變,但是也可以使個體具有某些優勢,例如帶有一個鐮刀型紅血球疾病基因的人,對瘧疾更有抵抗力。
對生物個體無益也無害的突變稱為中性突變,在族群中的出現頻率主要受到突變機率影響。由於這些突變不影響個體的生存機會,因此大多數物種的基因組在沒有天擇的狀況下,依然會有穩定數量的的中性突變不斷發生。單一鹼基對的變換稱為點突變,當一個或多個鹼基對插入或是刪除時,通常會使基因失去作用。
轉座子(transposon)是生物的基因組片段,並且在基因組的演化上扮演重要角色。它們能夠移動並插入基因組中,或是取代原有的基因,產生演化上的變異和多樣性。DNA複製也被認為每百萬年間,會在動物的基因組中產生數十到數百的新基因。
重組
在無性生殖的過程中,染色體上的任何一對等位基因都會一起遺傳到下一代。但是對於行有性生殖的物種而言,親代同源染色體中的等位基因,在製造生殖細胞的減數分裂過程中,會發生基因重組。這是一種不同的脫氧核糖核酸段落斷裂並重新組合的過程。
原核生物之間能夠透過接合等方式,直接交換彼此的基因,因此重組在原核生物中也比較常見。而較複雜的動物與植物,則通常是在製造生殖細胞的減數分裂時期,因為染色體的交接(crossover)而發生重組。減數分裂重組的發生頻率較低,而且排列位置較接近的等位基因,也較不易交換。因此可以由等位基因的重組率計算出基因的相對位置。
此外有性生殖中的孟德爾遺傳規則,能夠使有害的突變被清除,有益的突變被保留。且因為具有這種清除有害突變效果,因此當一個等位基因無法進行基因重組的時候(例如孤立的Y染色體),便因為有害突變逐漸累積,而使族群的有效族群大小(英語:effectivepopulationsize、簡寫:Ne)縮減,這種現象稱為希爾—羅伯森效應(Hill-Robertsoneffect)。若是染色體逐漸退化,則稱為繆勒氏齒輪(Muller'sratchet),這種現象比較容易出現在無性生殖的生物中。
進化機制
遺傳變異一方面經由生殖而傳遞到下一個世代(被稱為垂直基因轉移),另一方面也可以透過水平基因轉移(英語:horizontalgenetransfer,簡寫:HGT),在物種之內或是物種之間傳遞。尤其是細菌經常使用這種方式交換基因,最近的研究更發現可能有跨物種的水平基因轉移存在。基因流(geneflow)則是指基因在生物個體之間轉移。
基因型(遺傳因子)是產生表現型(外在表現)的根本。而表現型本身也擁有表型可塑性(phenotypicplasticity),能夠在基因型未改變的狀況下有所變化,並且能夠遺傳到下一世代。除了基因本身的改變,染色體的重新排列雖然不能改變基因,但是能夠產生生殖隔離,並使新物種形成。
一般來說,選擇包括了“天擇”(自然選擇)與“性擇”(性選擇)。天擇的主要原因是物種所居住環境的改變,包括物種之間關係的變化;性擇則是物種在繁殖的需求下而產生的選擇。而這些性擇所留下的性狀,可能會有害於個體本身的生存能力。各種選擇的分類事實上並不明確,也有一些分類以天擇表示所有選擇作用,並分為生態選擇(ecologicalselection)與性擇。
基因流
基因流也稱為遷移(migration),當族群之間並未受到地理或是文化上的阻礙時,基因變異會經由一些個體的遷移,使基因在不同族群間擴散,這樣的情形稱為基因流。恩斯特•麥爾認為基因流類似一種均質化(homogenising)的過程,因此能夠抵銷選擇適應的作用。當基因流受到某種阻礙,例如染色體的數目或是地理的隔閡,便會產生生殖隔離,這是物種形成的條件之一。
族群中等位基因的自由移動,也受到族群結構的阻礙,例如族群的大小或是地理分布。雖然理想狀態中族群的生殖對象完全自由且完全隨機,但是現實世界中並非如此,因此地理上的親近程度會對這些基因的移動造成龐大的影響。而且當遷移數量較少的時候,基因流對演化的影響也較低。
遺傳漂變
基因漂變指的是族群中等位基因頻率在每一個世代之間的隨機的變化。這種變化能夠以數學表達,哈蒂-溫伯格平衡描述了理想狀態情況下(不考慮天擇等因素)的數學模型。在理想狀態中,後代的等位基因頻率將接近隨機分布。當族群規模較大,基因漂變的機率會較低;當族群規模較小的時候,基因漂變的現象較為明顯。
當一個少數族群從原先族群之中分離而出,且兩者的基因頻率有所不同,若是分離而出的少數族群與原先族群的基因無法繼續交流,則兩者的基因頻率將漸行漸遠。這種現象稱為奠基者效應。例如從德國遷移到美國賓夕法尼亞的阿米什人,起源大約僅有200人,且習慣族內通婚。這個族群的埃利偉氏綜合症(Ellis-vanCreveldsyndrome)出現頻率較其它族群高。
水平基因轉移
以16SrRNA的基因序列所建立的種系發生樹,將生物演化歷程分為三域系統,包括細菌、古菌與真核生物。第一個提出這種分類的是卡爾•霍斯(CarlWoese)。由於水平基因轉移的存在,使生物的親源關係可能比圖片中所顯示的更為複雜許多。
水平基因轉移(英語:horizontalgenetransfer,簡寫:HGT),是個體將遺傳物質傳遞到其它非本身後代個體的過程。這種機制使遺傳物質得以在無直系關係的個體之間產生基因流。
水平基因轉移也可以經由抗原轉移(antigenicshift)、基因重整(reassortment)與雜交反應(hybridisation)等現象觀察。病毒能夠透過轉導作用(transduction)在物種間傳遞基因。細菌則能夠與死亡的細菌合體、經由轉形作用(transformation),以及與活細菌進行接合(conjugation),而獲得新的基因。而新的基因則能夠以質體的形式,加入宿主細菌的基因組中。雜交的現象在植物中最顯著,此外目前已知還有10種以上的鳥類物種能夠雜交。另外在哺乳動物與昆蟲中,也有雜交的例子,只是通常雜交後代不具有生殖能力。HGT也是細菌傳遞抗藥性的方式之一,而且有些發現表明HGT是原核生物與真核生物的演化重要機制。
由於HGT的存在,使種系發生學更加複雜,也使早期物種的演化過程出現一種隱藏關係(metaphor)。遺傳資訊在生殖作用之外,也能在物種之間傳遞。這使科學家必須在解釋演化關係的時候,表達出物種的隱藏關係,並且將不同的演化歷程組合.
自然選擇
 孔雀花枝招展的尾羽,是性擇的代表性例證,一方面使它容易成為被獵食的目標,另一方面又能夠吸引雌性。
孔雀花枝招展的尾羽,是性擇的代表性例證,一方面使它容易成為被獵食的目標,另一方面又能夠吸引雌性。由於各種基因的變異,使同一個族群中,不同個體的生存方式和繁殖方式有所不同,當環境發生改變,便會產生天擇作用。之所以稱為天擇,是因為這種選擇並非如基因漂變或基因突變一樣隨機,當環境改變發生時,將只有某些帶有特定特徵的群體能夠通過這些考驗。天擇有一些特例,有時候被視為與天擇擁有相等地位的選擇方式。其中包括性擇、人擇等等。
性擇指某個個體因為比起其它個體擁有較高的繁殖機會,因此它們的基因會被保留,使後代繼續保有相同的優勢。人擇指人類為了本身的生存或是喜好而對不同的基因變異進行篩選,通常發生在農業、畜牧業或是寵物的育種上。此外,優生學則是人類對人類所進行的篩選行為。不過人類事實上只是自然界的一部分,因此人擇與並天擇沒有明確的分別。
由於這些特殊的選擇機制,導致對於生物適應環境有益的特徵,並非在演化過程中一定會出現或是被保留。舉例而言,擁有更多的手指對人類的生活可能會更加方便,但是這種方便幾乎不會增加任何繁殖機會,甚至反而會減少。
演化思想史
早在古希臘時代,類似演化的思想已經出現,例如阿那克西曼德認為人類祖先來自海中。到了18世紀與19世紀,就已經有許多關於生命起源來自共同祖先的觀念。建立生物學的拉馬克,是第一位為演化提出科學理論的科學家。而達爾文與華萊士所提出,以天擇為主要機制的演化論,成為第一個具有說服力的解釋。在達爾文發表著作並成名之後,有許多先前尚未發表,發展較少的類似理論被發現。此外,達爾文在當時還未知任何遺傳機制,因此他無法解釋為何不同世代具有不同特徵。尤其當時流行子代的性狀為兩位親代性狀混合的概念,使得任何變異理論上似乎會逐漸消失。
孟德爾發現遺傳性狀的分離現象,解決了原本性狀混合的難題,然而一開始卻受到忽略。而且原本遺傳學家並不接受達爾文的天擇觀念,而是以突變作為演化原動力。直至20世紀,數學家羅納德•費雪、生物學家萊特(SewallWright)與霍爾登(J.B.S.Haldane)才建立了群體遺傳學,並與演化論結合。再加上湯瑪斯•摩根、特奧多修斯•多布然斯基、朱利安•赫胥黎、恩斯特•麥爾、喬治•辛普森、斯特賓斯(G.L.Stebbins)等人的研究,又稱為新達爾文主義的現代綜合理論在1920年到1940年代開始成形。
至於詳細的遺傳機制,則要等到埃弗里發現核酸為遺傳物質,以及華生與克里克,根據富蘭克林(RosalindFranklin)的研究,發表脫氧核糖核酸雙螺鏇結構,以及分子生物學的建立之後。
到了1960年代,許多生物學家開始以基因中心演化觀點探討演化過程。道金斯更認為,基因是唯一的天擇單位。此外還有漢彌爾頓提出以利他行為為基礎的親屬選擇。與之同時,古爾德與艾崔奇對演化的速率重新詮釋,提出疾變平衡論,認為生物的演化速度是長期的停滯與短暫的爆發所組合。之後演化生物學成形,並且發展出許多分支。
現今的研究與套用
演化生物學是研究演化的主要學門,探討物種的起源和改變,以及物種之間的親緣關係。這些研究影響了傳統的分類學,並導致系統分類學的出現。演化發育生物學(evo-devo)比較不同動物在發育過程中的變化,由此探討它們之間的關係與演化過程。體質人類學專注於人類的起源與演化,並探討人種的差異,又稱為生物人類學。
為了更深入研究演化過程與機制的細節,許多相關的分支學門產生。例如生態遺傳、人類演化、分子演化與種系發生。由於生物學是奠基在其它更基礎的自然科學之上,因此數學、統計學、物理學與化學對於了解演化機制也相當重要。例如為基因流、基因漂變等現象提供數學模式的群體遺傳學,研究在演化動力影響下,等位基因的分布和改變。
遺傳算法則是套用演化與遺傳的各種機制,並結合電腦的運算能力來解決許多問題。套用的層面包含工程、設計與通訊科技等。
演化在社會學的發展
有些演化研究專注在社會性生物上,稱為社會演化學。例如漢彌爾頓提出親屬選擇,解釋利他行為與邪惡的存在。不久之後,艾德華•威爾森出版的《社會生物學:新綜合》,解釋了社會性生物的各種行為,並在最後討論套用在人類行為的可能性。
1976年,道金斯在《自私的基因》一書中,認為人類的文化也能以演化解釋。他根據基因(gene)這個詞,將文化的演化單位稱為迷因(meme、在中文也被音譯為迷米)。類似作為遺傳因子的基因,迷因為文化的遺傳因子,也經由複製(模仿)、變異與選擇的過程而演化。
社會演化學與迷因學的差異在於,社會演化仍然是一種基因中心觀點,以遺傳物質分子為天擇單位;而迷因學則是以非基因的文化為天擇單位。
爭議
理論與事實
有時候演化會被人們強調為理論、學說或假說,而非真實。例如美國前總統里根曾在1980年的一場競選活動中表示:“演化是一個理論,只是一個科學理論,直到現在依然在科學界中受到挑戰,並且尚未被科學界認為絕對正確。”這類說法強調演化只是一個理論,所以並不是真實存在的事物。然而對科學家而言,理論並非與真實對立。真實是指經驗上所得的資料或數據,理論則是對事實的解釋與想法。
進步、複雜化與退化
有些物種(如人類),常被認為是比其它的物種更高級,甚至是演化的方向與目的所在。且認為演化的過程必定會使生物愈來愈複雜,或是進行與演化相反的退化。而現在的生物學家認為演化是沒有方向的過程,也沒有任何預先計畫的目標。雖然在已知的演化過程中,確實具有逐漸複雜的現象,但是依然有許多物種保持在較簡單的狀態,如細菌。因此複雜性可能增加也可能減少,或是維持不變,結果取決於天擇的機制。
物種形成
物種形成有時後被認為是無法直接觀察的現象,並得出演化是不科學的結論。但是科學的發現不僅是經由可重複的實驗,均變說(uniformitarianism)使科學家得以用經驗來推論事物的原因。此外物種形成的例子也出現在植物。還有刺魚(stickleback)的外胚葉發育不全(ectodysplasin)等位基因,被用來當作研究基因轉變與物種形成的模型。有一種類似的觀點,認為微觀演化是可以觀察,而巨觀演化則無法觀察。但是由於巨觀演化的機制與微觀演化相同,所以巨觀演化事實上已經在微觀演化中被觀察。而且物種之間基因序列的比較,也顯示少量的遺傳變異,就可以導致外表相當大的變化。
熵與生命
有些觀點認為演化增加複雜性的情形,違反了熱力學第二定律。熵是物理學上的亂度,這個定律是指在一個孤立系統中,熵只會增加或是維持不變,可以使用的自由能逐漸減少,最終反應逐漸趨於動態平衡。這種觀點忽略了生態系事實上並非孤立系統,所有生態系中生物所獲得的自由能,都是來自太空中,尤其是太陽。太陽、地球與太空的系統並不違反熱力學第二定律,因為太陽與地球輻射所產生的自由能,遠超過生物演化所需。
演化結果
適應
在天擇的作用影響之後,生物能夠更加適應它們所處的環境。只要是能夠使個體擁有更大生存優勢的過程,都可以稱為適應。不過需要注意的是,適應並非放諸四海皆準,在一個環境中擁有優勢的特徵,可能會在另一個環境中成為缺陷,這種現象也能解釋為何演化並沒有任何默認方向和目的,只有適應或是不適應。
即使“進步”在演化過程中並非必要,但是物種之間的競爭關係,仍能使物種在最基本的環境適應之外,進行更進一步的變化。這些競爭類似人類的軍備競賽(armsrace),且能夠依照物種的關係而分成兩類。一種是不對稱競爭,指物種受到不同條件的選擇,如掠食者與獵物的關係;另一種是對稱競爭,指物種受到相同的條件選擇,如森林中樹木對陽光的爭取。此外性擇也具有競爭的特性,有些物種在繁殖機會的競爭壓力之下,會逐漸發展出相當奇特的外觀或行為,例如鮮艷羽毛與吞食異性。
物種形成
物種形成受到許多類型的機制影響,主要可以分成2種類型。一種是異域性物種形成(allopatricspeciation),發生在族群受到地理隔離,而形成新物種的情況。例如大峽谷兩側的松鼠,由於峽谷地形的阻隔,形成兩個物種。不過地理隔離不一併會使物種分化,當兩個物種再度接近,有可能會產生雜交。例如台灣的白頭翁與烏頭翁,原本受到山脈阻隔而發展出不同型態,近年來純種烏頭翁卻逐漸與白頭翁交配而減少數量。同域性物種形成(sympatricspeciation),是指新物種與固有物種在相同的地區生存,因為繁殖或是溝通等行為而產生生殖隔離。例如印尼的一種蝙蝠由於聲音的不同,產生三個從未雜交的族群。
另外還有兩種較特殊的物種形成方式。邊域性物種形成(peripatricspeciation)介於異域性與同域性之間,指物種原先受地理隔離,之後因族群擴大而與固有物種接觸,卻已經形成兩個物種。邊域性物種形成是由恩斯特·麥爾所提出,這種理論同時支持了疾變平衡理論,且與奠基者效應有關,是小族群作為演化關鍵的例子。臨域性物種形成(parapatricspeciation)指物種雖然生存在相連的區域,但是因為交流的困難而產生新物種。最著名的現象是環狀物種(ringspecies),例如北極圈周圍的黑脊鷗(Larusargentatus)。
滅絕
 最知名的已滅絕動物之一,渡渡鳥
最知名的已滅絕動物之一,渡渡鳥滅絕指物種或是某個分類上的族群消失,並減少生物多樣性。某一物種的最後個體死亡,就是物種滅絕的時刻,即使滅絕前就已經失去了任何繁殖的可能。由於物種的潛在範圍可能相當大,因此確定物種滅絕時刻相當困難。
地球上曾經有過多次大規模的滅絕,其原因大多是因為環境,尤其是氣候的大幅改變。其中最嚴重的5次,分別是奧陶紀後期(4億4千萬年前)、泥盆紀後期(3億6千萬年前)、二疊紀後期(2億5千萬年前)、三疊紀後期(2億1千萬年前)與白堊紀末期(6千5百萬年前)。其中二疊紀後期的二疊紀滅絕事件,大約95%的海洋生物與70%的陸地動物消失。白堊紀末期的白堊紀滅絕事件,則因為恐龍的滅絕而著名。
社會爭議
 達爾文開創了演化論。
達爾文開創了演化論。在美國,這種創造論與演化論的爭議已經進到法院與政治層面。例如2005年的美國賓州多佛學區案中,法官判決學校在2004年開始教授的智慧設計論違反法律。以及同年美國總統布希公開贊成學校同時教授智慧設計論與演化論。
另一種對於演化論的批評,對象則是社會達爾文主義,以及衍生而出的種族主義、優生學與生育控制等19世紀末與20世紀初的產物。這類思想主要是起源於哲學家與政治人物對達爾文主義的借用,例如最早提出類似理論的赫伯特•史賓賽。而演化學者中也有一些本身具有優生學與種族主義等思想,例如提出胚胎重演論的海克爾。
