
脫氧核糖核酸重複順序
正文
在基因組脫氧核糖核酸中重複出現的核苷酸順序。與此相對,只出現一次或少數幾次的核苷酸順序稱為獨特順序或非重複順序。凡是比真菌複雜的真核生物都含有數量可觀的重複順序,在原核生物中則尚未發現。研究重複順序對了解生物進化和基因調控都有重要意義。復性動力學 美國學者R.J.布里頓等在1968年首先通過 DNA分子的復性動力學研究證實了真核生物細胞中存在著重複順序。加熱可以使 DNA雙鏈因鹼基對間的氫鍵斷裂而變為兩條獨立的單鏈,這一過程稱為變性。逐漸降溫又可以使兩條互補的單鏈恢復成為雙鏈,這一過程稱為復性或退火。復性過程是二級反應,反應的速度取決於兩個互補的單鏈片段相遇的機會,也就是取決於單鏈的濃度和反應時間。如果以復性的百分比為縱坐標,以反應時間(t,秒)和反應起始時的DNA單鏈的濃度(C0,核苷酸的克分子濃度)的乘積〔C0t,(克分子·秒)/升〕的對數為橫坐標作圖,則不同生物的DNA的復性進程常不相同(圖1);
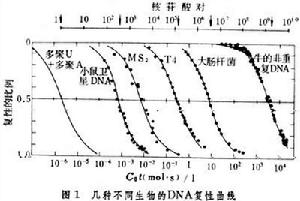 總的趨勢是生物的基因組越大,完成復性反應所需要的C0t值也愈大。復性達一半的 C0t值用C0t1/2表示。在特定的反應條件下,不同DNA有不同的C0t1/2值。它可以作為基因組的大小和順序複雜性的表征。
總的趨勢是生物的基因組越大,完成復性反應所需要的C0t值也愈大。復性達一半的 C0t值用C0t1/2表示。在特定的反應條件下,不同DNA有不同的C0t1/2值。它可以作為基因組的大小和順序複雜性的表征。 真核生物的DNA復性曲線通常不是一條簡單的S型曲線,而是可以分解成為若干部分的複雜的曲線,說明它們的DNA順序的複雜性。例如人的DNA復性曲線(圖2)
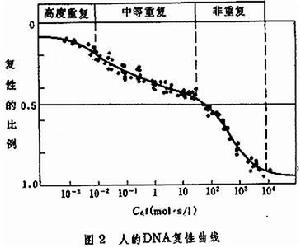 可以分解成為三個部分,分別代表高度重複順序、中等重複順序和非重複順序。
可以分解成為三個部分,分別代表高度重複順序、中等重複順序和非重複順序。 類型 各種不同類型的重複順序的差別表現在三個方面:①每種重複順序在基因組中出現的頻率;②組成每個重複單位的核苷酸數;③每種重複順序的特定的核苷酸順序。 依照復性反應的 C0t值,可以將重複順序分為高度重複順序和中等重複順序。前者的C0t1/2值一般在10-4~10-1之間,重複頻率大於105,它們通常由簡單的核苷酸順序組成,每個重複單位的長度大約是10~300個核苷酸對。某些生物的細胞核內的 DNA經氯化銫密度梯度超離心後,在主帶以外出現的一條衛星帶,這是一部分鹼基組成特殊而且高度重複的DNA,稱為衛星DNA。衛星DNA是串聯重複,在小鼠細胞總DNA中約占10%。衛星DNA往往分布在染色體著絲粒周圍和端粒的異染色質區中。在顯帶染色中含衛星DNA的部位以C帶的形式出現(見核型)。中等重複順序的C0t1/2值一般在10-1~102之間,重複頻率在102~105之間。 大部分中等重複順序與基因組中的獨特順序相間排列,散布在整個基因組中,重複單位的一般長度大約是300±200個核苷酸對。許多中等重複順序在細胞核中有它的相應的轉錄產物。至少有一部分這樣的順序屬於有功能的基因,例如編碼五種組蛋白的基因、核糖體RNA(rRNA)基因、5SRNA基因和各種轉運RNA(tRNA)基因,這些基因在基因組中的重複頻率往往在100~1000之間甚至更多。 它們在染色體上往往成串排列,形成基因簇。另一種特殊形式的重複順序是倒置重複順序,也稱回文順序。這是順序相同而取向相反的重複順序,C0t1/2值一般在10-5~10-4之間,變性後再復性時能在同一單鏈內形成“髮夾”式構型。
彼此間能夠復性的重複單位的核苷酸順序往往並不完全相同,這樣一組重複順序形成一個重複家族。實際上在任何一種生物中重複順序都以重複家族的形式存在。在每種生物中一般都有若干重複家族;在相近的物種中存在著類似的重複家族。例如在人的基因組中有一個Alu重複家族(根據它們都有共同的限制酶 Alu的切點而命名)每個成員的長度大約是300個核苷酸對。Alu家族散布在整個基因組中。現在已經發現在靈長類以及其他哺乳動物中都有類似 Alu的重複家族。在這方面的研究將為基因組的進化提供重要的線索。
功能 除了有已知基因功能的重複順序外,有些重複順序可以和核內不均一RNA中的成分進行分子雜交,這說明它們能夠轉錄。但迄今還不能判明它們在細胞生命活動中的確切功能。此外還有一部分重複順序,如衛星DNA則完全沒有轉錄產物,它們顯然不屬於結構基因。
目前對各種沒有一般基因功能的重複順序的生物學意義有幾種推測:①一部分重複順序可能是毫無意義的順序,它們在基因組中隨同其他DNA順序一起複製,是基因組中的“寄生物”,或稱“自私DNA”。②像衛星DNA這樣的高度重複順序可能參與染色體結構的維持,也可能與染色體配對和分離等行為有關,因此被稱為“家務DNA”; ③一些散布的重複順序可能作為基因間的間隔,組成所謂間隔 DNA;④部分中等重複順序可能參與基因調控,因為曾經發現在動物個體發育的不同時期以及不同組織中所轉錄的重複順序不同;⑤有些重複順序在較長的進化過程中可能作為新基因的素材,因為重複順序家族的成員似乎總是處在連續不斷的變化之中。
起源 布里頓於1968年提出突變複製假說,認為用進化的時間表來衡量,某種DNA順序的倍增是一種突發的事件,重複順序傳給後代,並可能通過自然選擇在種群中擴散。美國學者G.P.史密斯在1973年提出交換固定假說,它的基本前提是在姊妹染色單體間的不對等交換。他通過電子計算機模擬連續的交換過程,說明這種交換確實能導致重複順序的形成。這兩個假說都是推理性的,而且只能說明串接式重複順序的出現。現在已有一些研究結果表明某些散布的重複順序與倒置的重複順序有類似於轉座子的結構,例如酵母的Ty1順序、果蠅的copia順序、FB順序和人的Alu順序等,它們在基因組中的散布可能與轉座子的轉移現象有關(見轉座因子)。
