概述
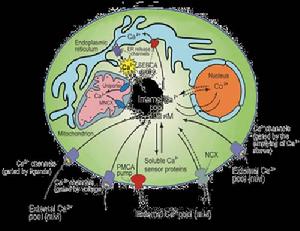 細胞鈣的分布及平衡
細胞鈣的分布及平衡鈣是生存與死亡信號,幾乎我們所有的生理活動都受到Ca2+ 的調控。
Ca2+ 作為細胞信使的基礎,是胞漿Ca2+ 與胞內鈣庫或胞外Ca2+ 之間存在濃度梯度。當某種刺激使胞內Ca2+ 濃度大幅度增加時,就起到傳遞信號的作用。 Ca2+ 本身的特性更有利於和靶蛋白結合,從而傳遞信息。由於要綜合多方面功能,Ca2+ 信號必然是非常靈活,同時又要受到嚴格的控制。它是藉助Ca2+ 信號的不同的中間環節來實現的,也就是Ca2+ 可以在不同的時間和空間上以不同幅度來起作用。不同類型的細胞以不同的鈣信號組合來完成其特定的生理功能。
鈣在細胞中的作用
Ca2+對細胞膜的興奮性是不可缺少的。當細胞內游離的Ca2+從靜息水平的10-8M上升時,則肌體的分泌、收縮、通道門控和其他過程都處於活動之中。鈣離子具有胞內信使作用。胞外鈣離子是胞內鈣離子的主要來源。鈣通道開放時,胞外Ca2+濃度的上升,可關閉電壓依賴性通道,增加靜息膜阻抗,從而也增加了神經和肌肉興奮性的閾值;若降低胞外Ca2+濃度則引起觸發性閾值變化,作用相反,神經和肌肉產生復極化,臨床上,病人表現為甲狀腺機能減退。
Ca2+是生理活性物質,在中樞神經系統中作用倍受人們關注。神經元不少關鍵的過程如:神經遞質的釋放、膜對特種離子的通透性的調節、膜的穩定性、軸漿流,以及許多酶的控制都是由細胞內或膜內、外鈣離子分布的改變來調節的
近年來的研究表明,肌細胞中肌漿網的Ca2+ATP酶的作用是將Ca2+從細胞內轉運並存到肌漿網內部。當Ca2+ATP酶的激活可以迅速降低胞液中Ca2+的濃度,讓肌肉得到舒張。心肌細胞內Ca2+濃度升高,可以引起心肌收縮。
紅細胞膜上的鈣泵,含有一個調節蛋白亞基,當細胞液中Ca2+濃度升高時,鈣調節蛋白的4個鈣離子結合位點與Ca2+結合,產生變構效應,激活Ca2+ATP酶,使Ca2+排到細胞外。鈣泵的作用,是維持細胞內Ca2+濃度,使之處於較低水平。
據報導:在非缺血情況下,興奮性胺基酸遞質結合突觸膜受體,引起Ca2+內流,其突觸反應的終止是通過興奮性胺基酸遞質被再攝取和Ca2+、Na+-K+ATP酶轉運系統使Ca2+從細胞內流出;在短暫性腦缺血時,使ATP耗竭,引起興奮性胺基酸再攝取和Ca2+外流減少,從而引起NMDA受體控制的Ca2+通道持續開放,結果細胞內Ca2+超載。
神經細胞鈣離子超載與老化有關。高濃度的Ca2+可以激活內質網上Ca2+,誘導Ca2+釋放通道,使內質網膜上釋放Ca2+。另外,一些激動劑可以與膜上受體結合,促發膜磷脂中磷醯肌醇-4,5二磷酸水解產生兩種新的肌內第二信使DAG還有IP3,調節神經細胞的活動。鈣超載對細胞損傷主要有幾個方面:
1. 酸化脫偶聯,呼吸作用受抑制;
2. 活磷脂酶C,A1,A2, 使膜磷脂水解,花生四烯酸(AA)釋放增加,同時產生大量自由基,使膜通透性增加,鈣離子大量流入,造成惡性循環;
3. 活依賴Ca2+的中性蛋白酶活性增加,造成神經元骨架破壞,蛋白質廣泛降解,神經細絲分解,神經微管解聚,嚴重影響軸漿運輸,導致神經元死亡。
Ca2+將死亡信號傳入細胞內的機制仍然知之甚少。當細胞內Ca2+濃度增加時,谷氨醯胺轉移酶活化,谷氨醯胺轉變成賴氨酸,從而形成位於膜下的穩定的交換蛋白,去參與凋亡小體的形成。Ca2+和鈣調蛋白能活化蛋白酶II,時細胞周期停止在G2,進而導致細胞凋亡。當細胞內Ca2+濃度降低,也能導致細胞凋亡,因為細胞內Ca2+濃度降低能抑制DNA和蛋白質合成,增加細胞內能量需求和影響許多Ca2+依賴性的細胞活動以及基因表達的變化等。
在許多組織中 ,細胞質Ca2+增加可活化細胞膜上的K+通道,使K+沿電化學梯度從胞內擴散到胞外。此外Ca2+還可以調節Ca2+、Cl- 和Na+通道的開啟,可以活化一類非專一性的陽離子通道。Ca2+對離子通道的活化與細胞的許多功能,特別是對細胞的興奮性傳導以及內分泌和外分泌均起著重要的調節作用。
近年研究發現,K+誘導的去極化可通過電壓門控Ca2+通道導致Ca2+的內流,而谷氨酸的作用則經突觸後非 NMDA受體介導引起Ca2+內流。視網膜中主要的膠質細胞受刺激後,會引起Ca2+從胞內鈣庫釋放後流至胞內不同部位,先是在細胞頂端Ca2+濃度升高,然後以波動的形式向終足區傳播。這種Ca2+波也許是視網膜中信號傳遞的第二條通路。
綜上所述,Ca2+在細胞功能的調節作用極大,在神經傳導、肌肉收縮、形態變化、神經細胞老化、腺體細胞的分泌和視網膜細胞的信號傳遞等生理過程都有Ca2+的調節參與。細胞的許多功能都依賴於細胞內外極高的Ca2+濃度差存在,一旦這種濃度差減低,細胞功能就會受到損傷,甚至引起細胞死亡。
胞內鈣離子的調控
胞漿內鈣離子來源於兩個方面,即近乎無限的外鈣內流和相對有限的內質網/肌漿網(ER/SR)內的鈣庫。外鈣進入途徑主要分成三大類:電壓門控的鈣通道(voltageoperatedcalciumchannel,VOC)、受體門控的鈣通道(receptoroperatedcalciumchannel,ROC)和鈣庫控制的鈣通道(storeoperatedcalciumchannel,SOC),VOC和ROC的產生機制相對研究的比較清楚,而SOC的產生機理卻仍然是一個謎。
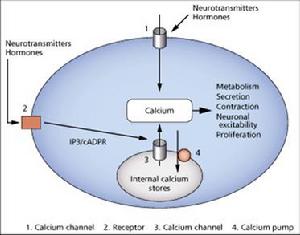 鈣泵及其受體
鈣泵及其受體SOC是受鈣庫的填充狀態來調節的鈣內流途徑,主要沒有解決問題涉及到鈣庫排空與通道開放之間的機制。VOC和ROC的特性是在短時間內產生大量的鈣進入,而SOC則產生較小的、然而卻是持續的鈣內流。胞記憶體在兩種鈣釋放通道,IP3和RYR(ryanodine)受體通道,將鈣離子從ER/SR鈣庫中釋放出來。由於細胞種類的差異性,RyR受體可以分成三種亞型,IP3受體則至少存在四種亞型。結構上兩種受體的C-末梢主域都有位於ER/SR上的跨膜區域,作為鈣釋放通道而存在著;而較大的N-末梢區域形成一個球根狀的頭部,伸到胞漿中去,整合控制通道開放和關閉的信號。另外,這兩種受體通道的開閉的控制機制也十分類似。由於它們的許多結構和生理上的相似之處,所以一般認為它們有相同的起源。細胞處於靜息狀態時,胞漿內的鈣離子通過複雜的機制來保持在穩定的低水平上。
 質膜鈣泵
質膜鈣泵通常,胞內的兩種鈣釋放通道的共同作用產生胞漿鈣離子的短暫上升從而實現鈣的信號作用。鈣離子本身也會以雙向的方式來影響通道的運行:當鈣離子濃度升高時,它會使受體對鈣離子的敏感性上升,增強通道的開放機率,促進鈣離子的進一步釋放,即產生鈣誘發的鈣釋放的過程(calciuminducedcalciumrelease,CICR);另一方面,一旦鈣離子濃度上升到一定程度時,它就會反過來抑制鈣離子的釋放,從而保證釋放出來的數量足以觸發生理功能而又不損傷細胞。如果這兩種受體僅僅只受鈣離子本身的調控,那么單個鈣釋放通道的隨機開放就可能觸發Ca上升而產生不需要的反應,因此這兩種受體必須還受到其它激動劑的控制。
對於IP3受體來說,另一種激動劑是IP3本身。綜合其他的研究表明,在強刺激產生了大量IP3情況下,它作為第二信使直接釋放鈣;而在生理條件下,IP3的上升雖然不足以直接釋放鈣但是它也會使IP3受體對鈣離子更加敏感來使得鈣信號可能發生。對於RyR受體說,也有類似的機制存在。環腺苷核酸核糖體(cADPR)會增強RyR受體的對鈣的敏感性,使胞漿成為可興奮性介質。總的來說,IP3和RyR型的鈣釋放通道的開閉都同時受到雙重激動劑的控制,協同作用產生複雜的鈣信號。
細胞膜構成了Ca2+流動的屏障,同時也是細胞功能調節的基礎。正常細胞中。膜內Ca2濃度比膜外Ca2+濃度低1萬倍左右,這並不等於說細胞內缺少Ca2+,事實上某些細胞器,如:線粒體、內質網和突軸小泡能攝取和貯存Ca2+,其中線粒體是細胞內最重要的鈣庫之一。另外,細胞內還有一些鈣結合到帶負電的脂和蛋白上,當細胞受刺激時,細胞外及細胞器中的Ca2+都可能被動的進入細胞質,使游離鈣濃度升至1~10 umol/L,從而引起一定的生理反應。
細胞內Ca2+濃度升高,主要由於Ca2+按濃度梯度通過Ca2+通道進入細胞的結果。膜系統上的Ca2+通道可以是電壓依賴的,也可以是激動劑依賴的,前者主要在肌肉和神經細胞中起作用。電壓依賴的鈣通道常有三種類型:
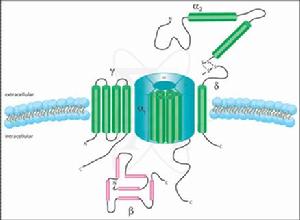 L型Ca2+通道的組成
L型Ca2+通道的組成L-型,長程型:需要較強的去激化刺激才能開放,失活也慢;
T-型,瞬時型:較弱的去激化電壓即能使通道開放,但失活也快;
N-型,需要較強的去激化電壓,失活快。
此外,神經元細胞上還存在一種P-型鈣通道,需要較強電壓激活,失活也慢。激動劑依賴型的鈣通道,也稱受體操縱性鈣通道,主要通過激動劑與質膜上特點受體結合後,啟動通道開放,使細胞外鈣進入細胞內,或使細胞器鈣庫釋放,使細胞內游離Ca2+上升。
細胞內Ca2+的排出是由另一類細胞功能蛋白——Ca2+泵來完成的。Ca2+泵逆濃度梯度將Ca2+排到細胞外或使其進入細胞內貯存庫中。細胞膜上還有另一類不直接與ATP水解偶聯的Ca2+主動傳遞,就是Na+/Ca2+交換體系,它是利用細胞膜兩側Na+濃度梯度通過Na+ -Ca2+交換將Ca2+排到細胞外。
可見,Ca2+在細胞內外保持動態平衡,依賴四個主要系統維持:
1、 電壓依賴性鈣通道;
2、 受體或神經遞質操控的鈣通道;
3、鈣泵;
4、細胞內第二信使(1、4、5三磷酸肌酸IP3)內在機制。
鈣通信的基本事件
近年來,由於光學成像技術的發展,如雷射共聚焦、雙光子激發、多光子激發等顯微技術的套用,使得實時觀測單個或是多個鈣釋放通道的活動成為可能[5,6]。這些通道的短暫開關會形成在空間範圍內的局部鈣升高(直徑大約為2μm),被稱為鈣信號(或是鈣通信)的基本事件。
它們的動力學特徵明顯,當通道打開時,鈣離子迅速擴散開來形成鈣梯度,呈現快速上升相;而當通道一旦關閉時,由於負反饋的作用,鈣離子擴散的速度較慢,並呈緩慢恢復相。目前,在細胞水平上,鈣信號可以分成三個層次;起初是最根本事件,來源於極低水平的刺激會導致單個通道的短暫開放,釋放出來的鈣被稱為鈣脈衝(calci-umblips)或鈣爆發(calciumquarks);然後是基本事件,它來源於一小群通道的開放,釋放出來的鈣形成鈣團(calciumpuffs)或鈣閃爍(calci2umsparks)。最後,在刺激強度較高時,大量的基本事件的同步化產生整體鈣信號。
基本事件有其特定的功能,但與整體鈣信號的功能不盡相同。在平滑肌細胞上發現,局部的鈣升高會作用於鈣離子敏感的K+通道,產生STOC,然後超極化細胞膜,從而使肌肉鬆弛。但如果鈣釋放位於細胞內部深處,使胞內鈣釋放通道共同作用則會產生了整體的的細胞鈣信號,肌肉就會收縮,這就是在同一個細胞內鈣信號的空間組織不同而激活完全相反反應的典型例子。一般來說,鈣信號的基本事件有兩個功能。基本事件影響靜息鈣水平:胞漿內的鈣水平是一個基本的鈣內流和清除的平衡。由於基本事件,如自發或誘發的wparks、puffs等,會釋放了一定量的鈣進入胞漿,因此它們就會影響胞漿的靜息游離鈣水平。
在細胞處於靜息狀態或受到刺激刺激程度很小時,每次基本事件中釋放出來的一定量的鈣對於胞內鈣水平施加了相當大的影響。這種鈣水平的提升可以增強胞內受體的興奮性,從而貢獻於整體的鈣信號。基本事件的局部信號功能:基本事件產生了空間上高度局限的鈣爆發,執行非常特定的信號功能。一個經典的例子是通過VOC的短暫鈣進入引發的細胞分泌過程。在這個過程中產生的基本事件雖然僅僅持續500μs,但是在某些分泌“熱點”(hotspot),局部鈣離子濃度會從100nm一直上升到200μm。因為VOC通過囊泡蛋白如聯接蛋白(syntaxin)和25kDa的突觸小體結合蛋白(synaptosome-associatedprotein,SNAP-25)與突觸囊泡接觸,所以基本事件應該位於錨定的囊泡的附近,這樣能夠用最短的時間來觸發分泌。
鈣離子通道的全局選擇性機制
鈣離子通道是一種跨越細胞膜的結構,它嚴格控制著鈣離子進入細胞的過程。由於鈣離子信號與很多重要生理功能相關,例如心臟收縮、基因轉錄等,因此調節鈣離子進入細胞的精確反饋機制就至關重要。為了實現這一功能,每個鈣離子通道都與一個鈣調蛋白分子(calmodulin CaM)結合,從而通過鈣離子與其羧基端小葉(C-lobe)和氨基端小葉(N-lobe)的結合實現對通道活性的調節。
鈣調蛋白與鈣離子形成的複合物是構成鈣離子感受器的重要原型,鈣離子感受器與鈣離子源密切相關。CaM的羧基端小葉能感應局域的鈣離子大幅振盪,這是由於主通道的鈣離子流引起的。而氨基端小葉則感應全局的較遠距離源引起的鈣離子小型改變。然而,儘管以上現象在生物學上非常重要,但其內部機制尚不清楚。
在2008年6月27日出版的《細胞》(Cell)上,來自美國的一組科學家發表文章稱,他們提出了一種全新理論來說明全局選擇性是如何出現的,並且從實驗上證實了這一理論的正確性。在研究中,科學家利用一種新方法實現了對於鈣離子振盪的毫秒級別控制。結果發現,全局選擇性產生於CaM結合於通道之後的快速鈣離子釋放。
儘管CaM的C-lobe和N-lobe感受著完全相同的鈣離子信號,它們卻選擇性的與產生於不同空間區域的鈣離子信號發生反應。研究人員發現,CaM的C-lobe利用一種“慢CaM”(slow CaM)機制來選擇產生於自身通道的鈣離子信號,這類似於放大鏡,而N-lobe則利用一種“SQS”機制來選擇來自較遠距離通道的信號,這類似於雙筒望遠鏡。特別值得注意的是,SQS機制產生的空間選擇性能得到調整,這或許能產生非常重要的生理學結果。
鈣調素
1968~1970年,美籍華人張槐耀在研究PDE對cAMP濃度的調節作用時,發現了鈣調素(CaM)。
 鈣調素的結構
鈣調素的結構一種能與鈣離子結合的蛋白質。鈣離子被稱為細胞內的第二信使,其濃度變化可調節細胞的功能,這種調節作用主要是通過鈣調素而實現的。
鈣調素是一種分子量為16700的單鏈蛋白質,由148個胺基酸組成。等電點為4.3,是酸性蛋白質。不同生物來源的鈣調素,其胺基酸組成和順序或完全一樣,或僅有少許差異。它耐酸,耐熱,十分穩定。鈣調素和細胞內很多種酶的作用有關。在每個鈣調素分子內,有4個可與鈣離子結合的區域,它們的一級結構極為相似。細胞內鈣離子水平通常維持在10-7摩爾濃度左右。當外來的刺激使細胞內鈣離子的濃度瞬息間升高至10-6~10-5摩爾濃度時,鈣調素即與鈣離子結合,構象改變,螺鏇度增加,成為活性分子,乃進而與酶結合,使之轉變成活性態。當鈣離子濃度低於10-6摩爾濃度時,鈣調素就不再與鈣離子結合,鈣調素和酶都復原為無活性態。因此,可以根據鈣離子濃度的變化來控制細胞內很多重要的生化反應。
至1982年,已得到電鰻和雞的鈣調素cDNA克隆和雞的鈣調素基因克隆。根據前二者的核苷酸順序分析,得知儘管它們的差異較大,但由之推測出的胺基酸順序仍基本相同。在雞的天然基因中,測得7個內含子,基因總長為12kb。在細胞分裂周期中,和細胞癌變時,鈣調素基因的表達加強。
CaM的結構特徵
①含30%的酸性胺基酸,為結合鈣提供羧基;
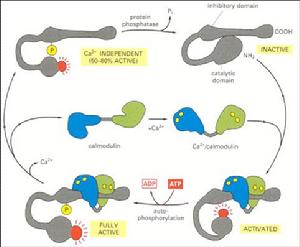 鈣調素的活化
鈣調素的活化②無易氧化的Cys和Trp,因而穩定、耐熱;
③不含能使肽鏈定型的hyp,因此具有高度靈活性,可與靶蛋白結合;
④ Phe和Tyr比值高,形成特異的有5個紫外吸收峰的光譜,可作為鑑定特徵。
⑤ 115位胺基酸為三甲基賴氨酸,可增加與靶酶的親和力。
鈣調素的活化及作用方式
CaM的作用方式:
①直接和靶酶結合,調節其活性;
②通過活化CaMK,再磷酸化靶酶,間接調節其活性。
CaM的活化:
CaM通過與Ca2+結合而活化。
CaM活化後的構象改變:①二級結構改變,導致疏水基團暴露,以利於與靶酶結合;②形成球形構象。
CaM激活靶酶的機制:
靶酶存在自抑制結構域,結合CaM後,消除了自抑制作用而被活化。
膜聯蛋白家族
與Ca2+結合後可進一步與膜磷脂結合的Ca2+結合蛋白,即鈣依賴性磷脂或膜結合蛋白。
此家族有12個成員,存在於細胞質膜下、儲Ca2+細胞器附近以及在核內或細胞外基質中。
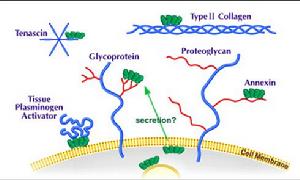 膜聯蛋白家族
膜聯蛋白家族膜聯蛋白家族的共同特點
1. 在mol/L濃度水平的Ca2+就可與磷脂或膜結合,而且對Ca2+及磷脂結合有特異性。
2. 在蛋白質的結構上有同源性。
3. 在細胞中的分布都直接或間接地與細胞質膜或內膜系統相聯繫。
膜聯蛋白的生理功能
參與胞吐作用;
在兩膜之間形成膜橋,有利於膜的黏著、聚集和融合;
具有離子通道功能;
對PLA2有抑制作用;
與細胞骨架有關。
