
概述
同位序列(Homeobox)或稱同源異型盒,是某些影響動物、真菌及植物發育的基因所擁有的一段DNA序列,擁有homeobox的基因稱作homeobox基因,統稱homeobox基因家族。
這段序列之長度約180個核苷酸,能轉錄出一段蛋白質區塊(protein domain),稱為homeobox區塊。
1995年9月9日,瑞典皇家科學院宣布,將1995年諾貝爾醫學獎授予三位在同位序列基因研究中做出突出貢獻的科學家:美國加利福尼亞理工學院的愛德華·劉易斯,美國普林頓大學的埃里克·維紹斯,德國馬克斯·普萊克發育生物研究所的女科學家克里斯蒂安·尼斯萊因·福爾哈德。
同位序列基因是在1984年被發現的,科學家們將這一發現譽之為“生物學中的一個劃時代的里程碑”。美國生物學家愛德華·劉易斯為了弄清生物的體型結構與基因之間的關係,進行了幾十年的研究,證實生物體從一單個的受精細胞開始,要發展成為含有數以億計的特化細胞的軀體,生物體內所有的細胞都含有一套完全相同的基因。但是,有些會變成骨骼的一部分,有些會變為抵抗疾病的抗體,而另一些則會長出毛髮。從蠅、蛙到人都擁有幾乎相同的一套基因。這套基因行使一個主控制功能,去引導動物的生長發育過程。
在胚胎生長過程中,每個細胞中的各種基因的開和關造成了細胞的特別功能。例如,產生眼球晶狀體蛋白的基因,只是在形成眼睛的細胞中才有活性;而創造腳趾甲蛋白的基因,即使它們都出現於同一個細胞中,卻不會被表達。
意義
同源異形盒模體在進化中廣泛存在。其重要意義是通過同源異形盒探針與很多真核的基因組進行雜交而獲得。在蛙,小鼠和人的DNA都發現同源異形盒的存在。在蛙和哺乳動物基因在胚胎髮生的早期表達增強了與果蠅一些基因的平行關係,表明可能這些會有同源異形的基因與各種生物胚胎髮育中基因調節有關。
詳細介紹
 同位序列
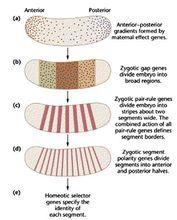
同位序列同位序列(homeobox)是同源異形基因中都具有的保守基序。控制果蠅發育的3組基因:母體基因,分節基因和同源異形基因。其中的一組調節另外兩個中的一組基因和編碼結構基因的靶基因。調節基因之間的相互作用是通過分析其它基因發生突變時相應基因的表達改變來確定的。但目前只鑑別出少數幾個結構靶基因,3組調節基因作用在靶上,使得身體的各部分產生分化。
分節基因編碼的蛋白調節轉錄,3個裂缺座位(hb,kr,KNI)基因含有鋅指模體。在轉錄因子TF和SP1中的模體是負責和DNA連線。裂缺基因中其它座位的產物也有DNA-結合模體;giant編碼的蛋白具有胺基酸拉鏈模體;而tailless編碼的蛋白與類固醇受體相似。這表明裂缺基因的一般功能是轉錄調節。在很多同源異形基因和分節基因中發現了保守的模體。普遍的模體是同位序列,它是位於各種分節基因和同源異形座位轉達錄單位3'端附近的180bp的區域。此序列是開放讀框的一部分。在這個序列中密碼子的第三個鹼基常發性改變。同位序列的大小及序列都是固定不變的。約有40個基因含有同位序列,而且這些基因幾乎都和發育調節有關(同位序列是通過它在同源異形基因中超優勢度(predominance)而首先被鑑別出來的,因此而得名)。
由同位序列編碼的蛋白序列稱之為同源結構域(homrodomain)。果蠅的同源結構域分成為幾組,主要是由BX-C/ANT-C復盒座位中同源異形基因組成。它們被稱為觸角足組。它們的同源結構域具有70-80%的保守性,通常是存在於蛋白質的C端。在相關的engrailed和invected基因中發現了特殊序列,它和觸角足組只有45%的同源性。其它類型的同源異形結構區序列存在於2~4個基因中。在果蠅中很多含有同位序列的基因組成了基因族。在BX-C基因簇中有3個同源異形基因具有同位序列。在ANT-C複合座位中含有一組5個帶有同位序列的同源異形基因,和4個也會有同位序列的其它基因。在BX-C和ANT-C中的同源異形基因有時也用HOM-C(homeotic-complex)基因來表示。在前-後軸上HOM-C的功能是什麼呢?據推測同源異形結構區上帶有不同的胺基酸序列可以識別DNA上不同的靶序列。
將不同的蛋白之間交換它們的同源異形結構區,這個實驗表明這些蛋白主要的特異部分就在乾同源異形結構區。但它們與特異的DNA靶位點的結合能力並不能表明它們的特點。例如,這些蛋白中的某一些在前後呼應中要么能激活轉錄,要么能抑制轉錄,它們的活性依賴於另一套能結合DNA,而卻不識別DNA的蛋白。在這一組基因中密切相關的一些成員的同源結構區之間的相似性表明它們能識別靶位點的重疊模式。這樣為組合效應開劈了一條途經,使帶有相關同源異形結構域的蛋白之間就能像依賴質量差異一樣依賴於數量,對同一位點,進行競爭。在有的情況下,不同的同源異形蛋白識別DNA上相同的靶位點,這也提出了有關它作用特異性的難題;估計DNA結合位點上仍存在著微小的差異或者存在著相互作用,例如蛋白質之間的相互作用。同位序列模體在進化中廣泛存在。其重要意義是通過同位序列探針與很多真核的基因組進行雜交而獲得。在蛙,小鼠和人的DNA都發現同位序列的存在。在蛙和哺乳動物基因在胚胎髮生的早期表達增強了與果蠅一些基因的平行關係,表明可能這些會有同源異形的基因與各種生物胚胎髮育中基因調節有關。在哺乳動物(可能所有動物)中那些與HOM-C組相關的基因都具有一個顯著的特點:它們像BX-C/ANT-C複合座位一樣組成基因簇。其中單個的哺乳動物的基因稱為Hox基因,其基因簇可達20-100kb,含有的基因可多達10個。4個Hox家族,基因含有同位序列,它們在小鼠和人類的基因組中被描敘。通過比較同位序列的序列表明哺乳動物中Hox基的排列和果蠅的同源基因是一致的。如HoxA4和HoxB4與Dfd最相關。
當哺乳動物的Hox基因簇和果蠅的同位序列基因簇的關係確定之後,人們就能識別一系列與ANT-C和BX-C基因簇相關的基因。哺乳動物Hox基因1~9組已被確定和ANT-C 和BX-C座位的序列相一致。10~13組是通過串聯重複產生的,與9組產生差異。在每一個基因簇中的相應座位有時稱為平行進化同源基因(paralogs),例如HoxA4和HoxB4就是平等進化同源基因。若果蠅和哺乳動物某一點上產生分歧的話,那么果蠅就可能存在只有單個的複合座位於,(如ANT-C和BX-C不同源)而複合座位含有的各基因都限定了前後極性。文昌魚(Amphioxus)其進化路線是和脊椎動物平行的。這種生物有單個的Hox基因簇,含有平行進化同源基因組中的一個成員;是原始基因簇的典型。在進化中果蠅的這些基因分成為兩個基因簇,而哺乳動 物整個一組基因發生了重複,在重複之後個別成員從各個複合座位中丟失了。這些基因在小鼠和果蠅中的排列和空間表達的方式都表現出平行性。在Hox基因簇中的基因在胚胎中表達方式是兩者在基因組中的組成呈對應的。這樣一來產生了一種很大的可能性,即基因簇不僅分擔了共同的進化,而且在基因組的組成與果蠅,小鼠空間表達的相關性中維持了相同的功能,在同源基因之間還存在著某些一致性。
小鼠的Hox D4 或HoxB6在果蠅中的異位表達產生的同源異形轉化與Dfd或Antp的同源異形表達的結果完全相同。從而證實對二者關係的推論。由於小鼠和果蠅蛋白之間的同源性,結果使它們保持了幾乎是它們所特有的異形結構域,這就加強了這樣的觀點,即結構域決定特異性。Hox基因很小,但具有大量的蛋白編碼單位。小鼠的HoxB基因簇長120kb 左右,含9個基因。基因組中的的位置和胚胎中表達兩者之間的連線與果蠅中的情況相類似,但前面只描敘了基因的本身;現在尚沒有掌握有關順式-作用位點的信息。這是因為在脊椎動物中通過各種突變來得出結論是十分困難的。但已發現控制Hox 基因僅通過上游區域的啟動子和增強子。這還要得看是否廣泛地存在著突變基因的副本,是否存在果蠅同源異形基因的複合控區。在果蠅中每個基因都是唯一的,但在脊椎動物中基因簇的重複使多基因(平等進化同源基因)具有相同或十分相似的表達模式。若平行進化同源基因有冗餘或部分的冗餘功能,那么缺乏一種產物至少可以被另一個基因簇相應基因產物所代替,因此突變的效應也就可減少到最小的程度。
小鼠中的Hox基因通過重排也可被破壞而導致隱性致死。在頭部和胸部的結構變異體中HoxA1和HoxA表達缺陷。不是在所有結構中突變基因都錯誤表達,表明確實存在功能的冗餘性。即1組或3組的Hox基因能彌補某些組織,缺乏HoxA基因而不是所有其它組織對HoxA基因的缺乏。在小鼠中同樣的突變能引起同源異形轉化,但要比果蠅中少。例如HoxC8的缺失使某些骨節出現前部骨節的表型。這還要看此是否是一般規律。Hox基因的異位表達已成功地用於論證,這些基因能使一些體節的特徵發生相應的轉化。最常見的是果蠅的前部體節出現了後部體節的表型。其實同源異型基因的表達是加上了附加信息。這些附加信息控制了後部體節的特徵。在小鼠中也獲得了相似的效應。但模式不完全一致。這些清楚表明Hox基因與果蠅決定前後軸發育模式的基因簇很相似。可能存在著Hox基因表達的組合密碼子或者可能在平行進化同源基因之間功能冗餘的程度不同。但目前尚未能提出它們在決定方式中所引起作用的模型。Hox基因座位結構仍難以解釋以什麼基因簇基因組的位置與胚胎的表達相一致?在進化中是如何維持下來的?一種解釋是基因表達的全面控制使得一個基因只有在基因簇中才能適當地表達。但這並不真實,至少在個別情況中一些基因可從基因簇中取消掉。
分析了啟動子區,表明Hox基因可受控於一系列的啟動子或增強子元件,啟動子和增強子控制了整個的表達模式。通常這些元件是在轉錄起始點的上游區域。例如HoxB4能在含有這些元件的質粒中表達,並可以製備轉基因鼠,但為什麼在進化壓力下在一個有序列的基因簇中保留著一些基因呢?一種可能是一個基因的增強子可能被埋入在另一個基因內。我們可以將一個基因易位別處時,看它是否有功能來加以判斷它是否屬於這種情況,若它含有相鄰基因的調節元件的話,那么它被取消將影響到其它基因的表達。
