
概念、特點
1.概念:有機物質在生物體細胞內氧化分解產生二氧化碳、水,並釋放出大量能量的過程稱為生物氧化(biological oxidation)。又稱細胞呼吸或組織呼吸。
2.特點:生物氧化和有機物質體外燃燒在化學本質上是相同的,遵循氧化還原反應的一般規律,所耗的氧量、最終產物和釋放的能量均相同。
(1)是在細胞內進行酶催化的氧化過程,反應條件溫和(水溶液中PH約為7和常溫)。
(2)在生物氧化的過程中,同時伴隨生物還原反應的產生。
(3)水是許多生物氧化反應的供氧體,通過加水脫氫作用直接參與了氧化反應。
(4)在生物氧化中,碳的氧化和氫化是非同步進行的。氧化過程中脫下來的質子和電子,通常由各種載體,如NADH等傳遞給氧並最終生成水。
(5)生物氧化是一個分步進行的過程。每一步都有特殊的酶催化,每一步反應的產物都可以分離出來。這種逐步反應的模式有利於在溫和的條件下釋放能量,提高能源利用率。
(6)生物氧化釋放的能量,通過與ATP合成相偶聯,轉換成生物體能夠直接利用的生物能ATP。
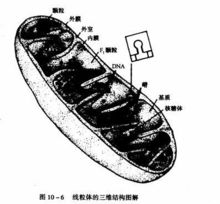
3.部位:在真核生物細胞內,生物氧化都是線上粒體內進行,原核生物則在細胞膜上進行。
氧化生成
 生物氧化
生物氧化生物氧化中CO2的生成是代謝中有機酸的脫羧反應所致。有直接脫羧和氧化脫羧兩種類型。按脫羧基的位置又有α-脫羧和β-脫羧之分。
中水的生成
(一)呼吸鏈的概念和類型
代謝物上的氫原子被脫氫酶激活脫落後,經過一系列的傳遞體,最後與激活的氧結合生成水的全部體系,此過程與細胞呼吸有關,所以將此傳遞鏈稱為呼吸鏈(respiratory chain)或電子傳遞鏈(electron transfer chain)。
在呼吸鏈中,酶和輔酶按一定順序排列線上粒體內膜上。其中傳遞氫的酶或輔酶稱為遞氫體,傳遞電子的酶或輔酶稱為電子傳遞體。遞氫體和電子傳遞體都起著傳遞電子的作用(2H→2H++2e)。
生物體內的呼吸鏈有多種型式。人體細胞線粒體內最重要的有兩條,即NADH氧化呼吸鏈和琥珀酸氧化呼吸鏈。它們的初始受氫體、生成ATP的數量及套用有差別。NADH氧化呼吸鏈套用最廣,糖、脂、蛋白質三大物質分解代謝中的脫氫氧化反應,絕大多數是通過該呼吸鏈來完成的。琥珀酸氧化呼吸鏈在Q處與上述NADH氧化呼吸鏈途徑交匯。其脫氫黃酶只能催化某些代謝物脫氫,不能催化NADH或NADPH脫氫。
(二)呼吸鏈的組成
組成呼吸鏈的成分已發現20餘種,分為5大類。
1.輔酶Ⅰ和輔酶Ⅱ
輔酶Ⅰ(NAD+或CoⅠ)為煙醯胺腺嘌呤二核苷酸。輔酶Ⅱ(NADP+或CoⅡ)為煙醯胺腺嘌呤二核苷酸磷酸。它們是不需氧脫氫酶的輔酶,分子中的煙醯胺部分,即維生素PP能可逆地加氫還原或脫氫氧化,是遞氫體。以NAD+作為輔酶的脫氫酶占多數。
2.黃素酶
黃素酶的種類很多,輔基有2種,即FMN和FAD。FMN是NADH脫氫酶的輔基,FAD是琥珀酸脫氫酶的輔基,都是以核黃素為中心構成的,其異咯嗪環上的第1位及第5位兩個氮原子能可逆地進行加氫和脫氫反應,為遞氫體。
3.鐵硫蛋白
分子中含有非血紅素鐵和對酸不穩定的硫,因而常簡寫為FeS形式。線上粒體內膜上,常與其他遞氫體或遞電子體構成複合物,複合物中的鐵硫蛋白是傳遞電子的反應中心,亦稱鐵硫中心,與蛋白質的結合是通過Fe與4個半胱氨酸的S相連線。
4.泛醌(又名輔酶Q)
一類廣泛分布於生物界的脂溶性醌類化合物。分子中的苯醌為接受和傳遞氫的核心,其C-6上帶有異戊二烯為單位構成的側鏈,在哺乳動物,這個長鏈為10個單位,故常以Q10表示。
5.細胞色素類
細胞色素(cytochrome, Cyt)是一類以鐵卟啉為輔基的結合蛋白質,存在於生物細胞內,因有顏色而得名。已發現的有30多種,按吸收光譜分a、b、c三類,每類又有好多種。
Cyta和a3 結合緊,迄今尚未分開,故寫成aa3,位於呼吸鏈的終末部位,其輔基為血紅素A,傳遞電子的機制是以輔基中鐵價的變化Fe3+ →Fe2+,a3還含有銅離子,把電子直接交給分子氧Cu+ →Cu2+,所以a3又稱細胞色素氧化酶。a3中的鐵原子可以與氧結合,也可以與氰化物離子(CN—)、CO等結合,這種結合一旦發生,a3便失去使氧還原的能力,電子傳遞中止,呼吸鏈阻斷,導致機體不能利用氧而窒息死亡。
(三)呼吸鏈中傳遞體的順序
呼吸鏈中氫和電子的傳遞有著嚴格的順序和方向。根據氧化還原原理,氧化-還原電勢E是物質對電子親和力的量度,電極電位的高低反映電子得失的傾向,E O'值愈低的氧還對(A/AH2)釋放電子的傾向愈大,愈容易成為還原劑而排在呼吸鏈的前面。所以NADH還原能力最強,氧分子的氧化能力最強。電子的自發流向是從電極電位低的物質(還原態)到電位高的氧化態,目前一致認可的是按標準氧還電位遞增值依次排列。
電子由NADH的傳遞到氧分子通過3個大的蛋白質複合體,即 NADH脫氫酶、細胞色素bc1複合體和細胞色素氧化酶到氧(又稱複合體Ⅰ、Ⅲ、Ⅳ)。電子從FADH2的傳遞是通過琥珀酸-輔酶Q還原酶(複合體Ⅱ)經Q、複合體Ⅲ、Ⅳ到氧(琥珀酸-輔酶Q還原酶催化的反應的自由能變化太小)。
氧化磷酸化
概念釋義
1.概念:氧化磷酸化(oxidative phosphorylation)是指在生物氧化中伴隨著ATP生成的作用。有代謝物連線的磷酸化和呼吸鏈連線的磷酸化兩種類型。即ATP生成方式有兩種。一種是代謝物脫氫後,分子內部能量重新分布,使無機磷酸酯化先形成一個高能中間代謝物,促使ADP變成ATP。這稱為底物水平磷酸化。如3-磷酸甘油醛氧化生成1,3-二磷酸甘油酸,再降解為3-磷酸甘油酸。另一種是在呼吸鏈電子傳遞過程中偶聯ATP的生成。生物體內95%的ATP來自這種方式。
偶聯部位
2.偶聯部位:根據實驗測定氧的消耗量與ATP的生成數之間的關係以及計算氧化還原反應中ΔGO'和電極電位差ΔE的關係可以證明。
P/O比值是指代謝物氧化時每消耗1摩爾氧原子所消耗的無機磷原子的摩爾數,即合成ATP的摩爾數。實驗表明, NADH在呼吸鏈被氧化為水時的P/O值約等於3,即生成3分子ATP?(1~6版生物化學此數據為3,7版為2.5);FADH2氧化的P/O值約等於2,即生成2分子ATP(1~6版生物化學此數據為2,7版為1.5)。
氧-還電勢沿呼吸鏈的變化是每一步自由能變化的量度。根據ΔGO'= - nFΔE O'(n是電子傳遞數,F是法拉第常數),從NADH到Q段電位差約0.36V,從Q到Cytc為0.21V,從aa3到分子氧為0.53V,計算出相應的ΔGO'分別為69.5、40.5、102.3kJ/mol。於是普遍認為下述3個部位就是電子傳遞鏈中產生ATP的部位。
NADH→NADH脫氫酶→‖Q → 細胞色素bc1複合體→‖Cytc →aa3→‖O2
氧化作用
糖代謝中的三羧酸循環和脂肪酸β-氧化是線上粒體內生成NADH(還原當量),可立即通過電子傳遞鏈進行氧化磷酸化。在細胞的胞漿中產生的NADH ,如糖酵解生成的NADH則要通過穿梭系統(shuttle system)使NADH的氫進入線粒體內膜氧化。
(一)α-磷酸甘油穿梭作用
這種作用主要存在於腦、骨骼肌中,載體是α-磷酸甘油。
胞液中的NADH在α-磷酸甘油脫氫酶的催化下,使磷酸二羥丙酮還原為α-磷酸甘油,後者通過線粒體內膜,並被內膜上的α-磷酸甘油脫氫酶(以FAD為輔基)催化重新生成磷酸二羥丙酮和FADH2,後者進入琥珀酸氧化呼吸鏈,生成1.5分子ATP。葡萄糖在這些組織中徹底氧化生成的ATP比其他組織要少,1摩爾G→30摩爾ATP。
(二)蘋果酸-天冬氨酸穿梭作用
主要存在肝和心肌中。1摩爾G→32摩爾ATP
胞液中的NADH在蘋果酸脫氫酶催化下,使草醯乙酸還原成蘋果酸,後者藉助內膜上的α-酮戊二酸載體進入線粒體,又線上粒體內蘋果酸脫氫酶的催化下重新生成草醯乙酸和NADH。NADH進入NADH氧化呼吸鏈,生成2.5分子ATP。草醯乙酸經穀草轉氨酶催化生成天冬氨酸,後者再經酸性胺基酸載體轉運出線粒體轉變成草醯乙酸。
偶在線上制
(一)化學滲透假說(chemiosmotic hypothesis)
1961年,英國學者Peter Mitchell提出化學滲透假說(1978年獲諾貝爾化學獎),說明了電子傳遞釋出的能量用於形成一種跨線粒體內膜的質子梯度(H+梯度),這種梯度驅動ATP的合成。這一過程概括如下:
1.NADH的氧化,其電子沿呼吸鏈的傳遞,造成H+ 被3個H+ 泵,即NADH脫氫酶、細胞色素bc1複合體和細胞色素氧化酶從線粒體基質跨過內膜泵入膜間隙。
2.H+ 泵出,在膜間隙產生一高的H+ 濃度,這不僅使膜外側的pH較內側低(形成pH梯度),而且使原有的外正內負的跨膜電位增高,由此形成的電化學質子梯度成為質子動力,是H+ 的化學梯度和膜電勢的總和。
3.H+ 通過ATP合酶流回到線粒體基質,質子動力驅動ATP合酶合成ATP。
(二)ATP合酶
ATP合酶由兩部分組成(Fo-F1),球狀的頭部F1突向基質液,水溶性。亞單位Fo埋在內膜的底部,是疏水性蛋白,構成H+ 通道。在生理條件下,H+ 只能從膜外側流向基質,通道的開關受柄部某種蛋白質的調節。
相關因素
(一)抑制劑
能阻斷呼吸鏈某一部位電子傳遞的物質稱為呼吸鏈抑制劑。
魚藤酮、安密妥在NADH脫氫酶處抑制電子傳遞,阻斷NADH的氧化,但FADH2的氧化仍然能進行。
抗黴素A抑制電子在細胞色素bc1複合體處的傳遞。
氰化物、CO、疊氮化物(N3-)抑制細胞色素氧化酶。
對電子傳遞及ADP磷酸化均有抑制作用的物質稱氧化磷酸化抑制劑,如寡黴素。
(二)解偶聯劑
2,4-二硝基苯酚(DNP)和頡氨黴素可解除氧化和磷酸化的偶聯過程,使電子傳遞照常進行而不生成ATP。DNP的作用機制是作為H+的載體將其運回線粒體內部,破壞質子梯度的形成。由電子傳遞產生的能量以熱被釋出。
(三)ADP的調節作用
正常機體氧化磷酸化的速率主要受ADP水平的調節,只有ADP被磷酸化形成ATP,電子才通過呼吸鏈流向氧。如果提供ADP,隨著ADP的濃度下降,電子傳遞進行,ATP在合成,但電子傳遞隨ADP濃度的下降而減緩。此過程稱為呼吸控制,這保證電子流只在需要ATP合成時發生。
