
概述
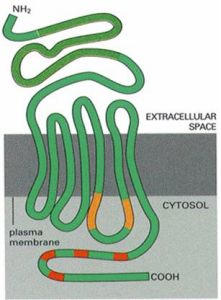 G蛋白耦聯型受體為7次跨膜蛋白
G蛋白耦聯型受體為7次跨膜蛋白三聚體GTP結合調節蛋白(trimericGTP-bindingregulatoryprotein)簡稱G蛋白,位於質膜胞質側,由α、β、γ三個亞基組成,α和γ亞基通過共價結合的脂肪酸鏈尾結合在膜上,G蛋白在信號轉導過程中起著分子開關的作用(圖8-12),當α亞基與GDP結合時處於關閉狀態,與GTP結合時處於開啟狀態,α亞基具有GTP酶活性,能催化所結合的ATP水解,恢復無活性的三聚體狀態,其GTP酶的活性能被rgs(regulatorofGproteinsignaling)增強。RGS也屬於GAP(GTPaseactivatingprotein)。
G蛋白耦聯型受體為7次跨膜蛋白,受體胞外結構域識別胞外信號分子並與之結合,胞內結構域與G蛋白耦聯。通過與G蛋白耦聯,調節相關酶活性,在細胞內產生第二信使,從而將胞外信號跨膜傳遞到胞內。G蛋白耦聯型受體包括多種神經遞質、肽類激素和趨化因子的受體,在味覺、視覺和嗅覺中接受外源理化因素的受體亦屬G蛋白耦聯型受體。
由G蛋白耦聯受體所介導的細胞信號通路主要包括:cAMP信號通路和磷脂醯肌醇信號通路。
CAMP信號途徑
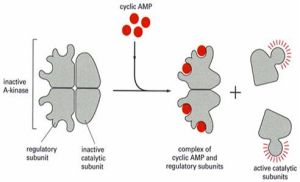 蛋白激酶A
蛋白激酶A在cAMP信號途徑中,細胞外信號與相應受體結合,調節腺苷酸環化酶活性,通過第二信使cAMP水平的變化,將細胞外信號轉變為細胞內信號。
1、cAMP信號的組分
①.激活型激素受體(Rs)或抑制型激素受體(Ri);
②.活化型調節蛋白(Gs)或抑制型調節蛋白(Gi);
③.腺苷酸環化酶(Adenylylcyclase):是相對分子量為150KD的糖蛋白,跨膜12次。在Mg2+或Mn2+的存在下,腺苷酸環化酶催化ATP生成cAMP。
④.蛋白激酶A(ProteinKinaseA,PKA):由兩個催化亞基和兩個調節亞基組成(圖8-15),在沒有cAMP時,以鈍化複合體形式存在。cAMP與調節亞基結合,改變調節亞基構象,使調節亞基和催化亞基解離,釋放出催化亞基。活化的蛋白激酶A催化亞基可使細胞內某些蛋白的絲氨酸或蘇氨酸殘基磷酸化,於是改變這些蛋白的活性,進一步影響到相關基因的表達。
⑤.環腺苷酸磷酸二酯酶(cAMPphosphodiesterase):可降解cAMP生成5’-AMP,起終止信號的作用。
磷脂醯肌醇途徑
磷脂醯肌醇途徑在磷脂醯肌醇信號通路中胞外信號分子與細胞表面G蛋白耦聯型受體結合,激活質膜上的磷脂酶C(PLC-β),使質膜上4,5-二磷酸磷脂醯肌醇(PIP2)水解成1,4,5-三磷酸肌醇(IP3)和二醯基甘油(DG)兩個第二信使,胞外信號轉換為胞內信號(圖8-21),這一信號系統又稱為“雙信使系統”(doublemessengersystem)。
IP3與內質網上的IP3配體門鈣通道結合,開啟鈣通道,使胞內Ca2+濃度升高。激活各類依賴鈣離子的蛋白。用Ca2+載體離子黴素(ionomycin)處理細胞會產生類似的結果。
DG結合於質膜上,可活化與質膜結合的蛋白激酶C(ProteinKinaseC,PKC)。PKC以非活性形式分布於細胞溶質中,當細胞接受刺激,產生IP3,使Ca2+濃度升高,PKC便轉位到質膜內表面,被DG活化(圖8-22),PKC可以使蛋白質的絲氨酸/蘇氨酸殘基磷酸化是不同的細胞產生不同的反應,如細胞分泌、肌肉收縮、細胞增殖和分化等。DG的作用可用佛波醇酯(phorbolester)模擬。
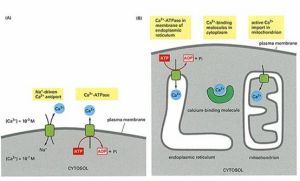 Ca2+信號的消除
Ca2+信號的消除Ca2+活化各種Ca2+結合蛋白引起細胞反應,鈣調素(calmodulin,CaM)由單一肽鏈構成,具有四個鈣離子結合部位。結合鈣離子發生構象改變,可激活鈣調素依賴性激酶(CaM-Kinase)。細胞對Ca2+的反應取決於細胞內鈣結合蛋白和鈣調素依賴性激酶的種類。如:在哺乳類腦神經元突觸處鈣調素依賴性激酶Ⅱ十分豐富,與記憶形成有關。該蛋白發生點突變的小鼠表現出明顯的記憶無能。
IP3信號的終止是通過去磷酸化形成IP2,或被磷酸化形成IP4。Ca2+由質膜上的Ca2+泵和Na+-Ca2+交換器將抽出細胞,或由內質網膜上的鈣泵抽進內質網。
DG通過兩種途徑終止其信使作用:一是被DG-激酶磷酸化成為磷脂酸,進入磷脂醯肌醇循環;二是被DG酯酶水解成單酯醯甘油。由於DG代謝周期很短,不可能長期維持PKC活性,而細胞增殖或分化行為的變化又要求PKC長期活性所產生的效應。現發現另一種DG生成途徑,即由磷脂酶催化質膜上的磷脂醯膽鹼斷裂產生的DG,用來維持PKC的長期效應。
其它G蛋白偶聯型受體
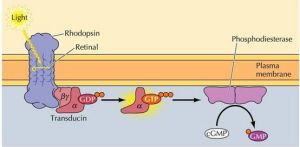 視覺感受器中的G蛋白
視覺感受器中的G蛋白1.化學感受器中的G蛋白
氣味分子與化學感受器中的G蛋白偶聯型受體結合,可激活腺苷酸環化酶,產生cAMP,開啟cAMP門控陽離子通道(cAMP-gatedcationchannel),引起鈉離子內流,膜去極化,產生神經衝動,最終形成嗅覺或味覺。
2.視覺感受器中的G蛋白
黑暗條件下視桿細胞(或視錐細胞)中cGMP濃度較高,cGMP門控鈉離子通道開放,鈉離子內流,引起膜去極化,突觸持續向次級神經元釋放遞質。
視紫紅質(rhodopsin,Rh)為7次跨膜蛋白,含一個11順-視黃醛。是視覺感受器中的G蛋白偶聯型受體,光照使Rh視黃醛的構象變為反式,Rh分解為視黃醛和視蛋白(opsin),構象改變的視蛋白激活G蛋白(transducin,Gt),G蛋白激活cGMP磷酸二酯酶,將細胞中的cGMP水解。從而關閉鈉通道,引起細胞超極化,產生視覺。可見胞內cGMP水平下降的負效應信號起傳遞光刺激的作用。
視覺感受器的換能反映可表述為:
光信號→Rh激活→Gt活化→cGMP磷酸二酯酶激活→胞內cGMP減少→Na+離子通道關閉→離子濃度下降→膜超極化→神經遞質釋放減少→視覺反應。
小G蛋白
小G蛋白(SmallGProtein)因分子量只有20~30KD而得名,同樣具有GTP酶活性,在多種細胞反應中具有開關作用。第一個被發現的小G蛋白是Ras,它是ras基因[5]的產物。其它的還有Rho,SEC4,YPT1等,微管蛋白β亞基也是一種小G蛋白。
小G蛋白的共同特點是,當結合了GTP時即成為活化形式,這時可作用於下游分子使之活化,而當GTP水解成為GDP時(自身為GTP酶)則回復到非活化狀態。這一點與Gα類似,但是小G蛋白的分子量明顯低於Gα。
在細胞中存在著一些專門控制小G蛋白活性的小G蛋白調節因子,有的可以增強小G蛋白的活性,如鳥苷酸交換因子(guaninenucleotideexchangefactor,GEF)和鳥苷酸解離抑制因子(GuaninenucleotidedissociationInhibitor,GDI),有的可以降低小G蛋白活性,如GTP酶活化蛋白(GTPaseactivatingprotein,GAP)。
