什麼是防禦素
防禦素是一類可殺死細菌、真菌或者病毒等微生物的多肽[1]。天然的防禦素分布在從植物、低等動物到哺乳動物等幾乎所有的生物類群中,是生物長期與疾病鬥爭中進化而來的,也是其自身防禦體系的重要組成部分。大部分防禦素和其他蛋白質類物質一樣,由基因直接編碼,可通過基因工程技術開發生產。
防禦素研究背景
1966年,美國科學家Zeya和Spitznagel首次在哺乳動物老鼠和豚鼠的多形核嗜中性白細胞中發現一類具有抗菌活性的鹼性多肽,將其稱之為“溶酶體陽離子蛋白”。這就是後來人們稱之為防禦素的物質。到目前,人們已經鑑定了四百餘種防禦素。
1985年,美國加州大學RobertLehrer博士首次命名防禦素(defensins),並報導了兔防禦素MCP-1及MCP-2對流感病毒和單純皰疹病毒具有直接中和滅活作用。次年,該研究組還報導了人體內三種alpha防禦素(Humanα-defensin或humanneutrophilpeptide,HNP)對流感病毒、巨細胞病毒、水泡性口炎病毒等均有中和作用。
2002年,美國洛克菲勒大學艾倫•戴蒙德愛滋病研究中心的華人科學家張林琦在《SCIENCE》雜誌上發表文章,報導人體內抑制愛滋病病毒複製的物質是T淋巴細胞分泌的α-防禦素,這個發現為愛滋病的控制與治療提供了新的思路與方向,引起生物醫學界的廣泛關注。
2003年開始,和仕康公司的核心研發人員在構建海洋生物海馬等物種的cDNA文庫時發現了大量防禦素基因,並分離到五種天然防禦素。2007年開始,他們依託南海海洋生物技術國家工程中心等科研轉化平台將防禦素髮酵製劑作為飼料添加劑進行大量動物實驗,結果表明該製劑可顯著提升動物免疫力和防病抗病水平。
防禦素的分類
(一)哺乳動物類防禦素
防禦素是動物體內防禦系統的主要成分,大多由29~42個胺基酸殘基組成,含3對分子內二硫鍵,相對分子質量為2~6ku,根據其二硫鍵位置的不同可分為α-防禦素、β-防禦素、θ-防禦素3類。
α-防禦素是於1980年美國Lehrer實驗室從兔肺巨噬細胞中首先分離得到的、陽離子性極強的小分子抗菌肽,稱為defensin,後被歸為α-防禦素;主要分布於人、兔、豬、鼠類的嗜中性粒細胞、兔子的齒槽巨嗜細胞及人類和齧齒動物的小腸潘氏細胞。α-防禦素分子鏈內二硫鍵的連線位置分別為Cys1-Cys6、Cys2-Cys4、Cys3-Cys5。其中Cys1-Cys6連線N端和C端,形成分子大環。
β-防禦素是Diamond等(1991)首先在牛的氣管黏膜上皮細胞中發現的,後又在牛粒性白細胞中發現了13種與其序列高度相似,但其共有序與α-防禦素不同,故被命名為β-防禦素;主要分布於牛的骨髓和人及多種動物(牛、羊、豬、駱駝、馴鹿、小鼠、大鼠)的胃腸道、呼吸道、舌、牙齦、腎、皮膚的上皮中,近來在梅花鹿舌黏膜的上皮細胞內也發現了該短肽。單核細胞和巨噬細胞通常缺乏防禦素,但是他們可以釋放誘導上皮細胞合成β-防禦素的信使。β-防禦素分子鏈內二硫鍵的連線位置分別為Cys1-Cys5、Cys2-Cys4、Cys3-Cys6。
θ-防禦素是於2002年Trabi等用反向高效液相層析色譜法從獼猴的白細胞中分離出來的1種成環狀結構分子,又被稱為獼猴θ型防禦素-1(RTD-1),主要分布於巨嗜細胞內。θ-防禦素的結構不同於α和β-防禦素,其前體(已經發現3種)是α-防禦素類似物,由1個終止密碼子從α-防禦素3個半胱氨酸碳骨架的第4個殘基處截斷,從截短的α-防禦素前體切掉1個9個胺基酸的片段,然後從頭到尾進行剪下,直到其他的相同或相似的九肽出現。成熟的θ-防禦素是2個半防禦素的修飾與結合的產物,它的前體(稱之為半防禦素)是變異的α-防禦素基因和1個未成熟的終止密碼子的編碼產物,導致每1個前體僅僅包含3個半胱氨酸殘基。θ-防禦素分子鏈內二硫鍵的連線位置分別為Cys1-Cys4、Cys2-Cys5、Cys3-Cys6,連線形成環狀結構。
(二)昆蟲類防禦素[2]
在1988年由Masturyama在1種半翅目昆蟲肉蠅中發現了第1種昆蟲防禦素。後來Dimarcq等先從果蠅中分離到1種病原菌誘導的抗菌肽,發現果蠅的防禦素基因是單拷貝且無內含子,據此推測昆蟲的防禦素是獨立進化而來的。後來經研究發現,昆蟲防禦素與哺乳動物防禦素在二硫鍵的連線方式以及三維空間構型上是截然不同的,從而證明了昆蟲防禦素和哺乳動物防禦素不具同源性。絕大多數昆蟲的防禦素帶1個淨正電荷,分子量為4ku左右,由34~51個胺基酸組成,胺基酸序列中都含有6個保守半胱氨酸,3個分子內二硫鍵形成β片狀結構和1個α螺鏇結構,二硫鍵以Cys1-Cys4、Cys2-Cys5、Cys3-Cys6連線。
(三)植物類防禦素
在1990年由Mendez等從小麥、大麥種子首先分離得到植物防禦素r1-P和r1-H,用NMR光譜法測定其三維結構,發現與哺乳動物和昆蟲防禦素相似。最近Thevissen也通過比較植物和昆蟲防禦素與真菌的葡糖苷鞘氨醇的相互作用,得知它們是由同一個前體細胞祖先進化而來的。植物防禦素的分子質量小於5ku,由45~54個胺基酸組成,3個分子內二硫鍵形成1個反向的β片狀結構和1個α螺鏇結構,含有4對二硫鍵且連線方式為Cys1-Cys8、Cys2-Cys5、Cys3-Cys6、Cys4-Cys7。
防禦素的生物活性
(一)抗菌作用
防禦素能有效殺滅革蘭陰性細菌和革蘭陽性細菌。在體外濃度為10~100mg/L的防禦素即對多種細菌具有殺傷作用,而防禦素在中性粒細胞中的濃度為g/L級,遠遠超過上述數值,這表明在體內防禦素可能具有更強的殺菌活性,目前研究發現防禦素對革蘭陽性細菌的殺傷能力明顯要強於革蘭陰性細菌。在體外,HBD-2對大腸桿菌的半數致死量(LD50)為0.46nmol/ml,最小抑菌濃度(MIC)為15μg/ml,而對綠膿桿菌、金黃色葡萄球菌的MIC為62μg/ml。體外實驗證明大多數防禦素的MIC範圍為0.5-10μmol/L[3]。
對於防禦素抗菌活性機制,大部分研究者認為主要和微生物的細胞膜結構有關。防禦素髮揮抗菌作用可分為三個階段:
(1)由靜電吸引。防禦素與靶細胞膜結合。防禦素帶正電荷,可通過靜電作用與帶負電荷的細菌膜脂層結合;
(2)通道形成。帶正電荷的防禦素分子或其多聚體則與細菌質膜上帶負電荷的磷脂頭部和水分子相互作用,顯著地增加生物膜的通透性。防禦素作用於膜上形成穩定的多個通道;
(3)內容物外泄。通道形成後,防禦素進入細胞內的同時,其他胞外分子也伴隨進入(如肽、蛋白質或無機離子),而靶細胞的重要物質(如鹽離子和大分子)滲出,致使靶細胞發生不可逆損傷而死亡。
防禦素還能通過誘導細胞因子的釋放和介導非成熟性的樹突狀細胞協同刺激分子的表達上調,促進IDC的成熟,進而活化T細胞,觸發特異性免疫應答。
(二)抗病毒作用
防禦素能殺滅一些被膜病毒,如HIV、皰疹病毒、水泡型口炎病毒,但對無衣殼病毒卻無效。θ-防禦素還具有抗濾過性病原體和抗毒素作用。體內實驗顯示,防禦素可延緩或根除兔梅毒病,並使兔牙周炎齦下菌群恢復正常。防禦素主要通過與病毒外殼蛋白結合從而導致病毒喪失生物活性,這一特殊作用機理也使得微生物不易對其產生抵抗性。防禦素可直接抑制病毒,對病毒的抑制程度依賴於防禦素濃度以及分子內二硫鍵的緊密程度,其抗病毒功效同樣受時間、pH值、離子強度和溫度等因素的影響,在中性及低離子強度條件下,防禦素具有較強的抗病毒活性[4],而在實驗體系中加入血清或血清蛋白則可大大削弱防禦素的抗病毒功效。有關防禦素的抗病毒作用機理,大體上可以總結為如下三點:
1、閉門羹----阻止病毒入侵宿主細胞
許多細胞和病毒的外膜分子是糖蛋白,它們就像刷子一樣突起(見圖一)。
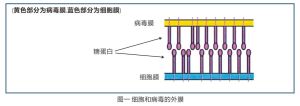 防禦素阻止病毒入侵宿主細胞
防禦素阻止病毒入侵宿主細胞 病毒感染細胞採用“兩步走”政策:首先,病毒的外衣即包膜死纏爛打地粘著到細胞的外膜上;接著,病毒包膜與細胞膜融合在一起。兩個膜融合後,病毒就將它的遺傳物質插入到細胞中。
防禦素則斜插在糖蛋白上以防病毒向細胞糖蛋白的擴散(見圖二),讓病毒吃個“閉門羹”無法進入細胞。沒能進入細胞的病毒接著會被免疫系統的細胞摧毀。
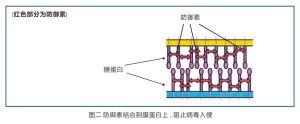 防禦素阻止病毒入侵宿主細胞
防禦素阻止病毒入侵宿主細胞 2、突破口----殺滅病毒
防禦素通常帶多個正的淨電荷,而病毒囊膜及其表面糖蛋白通常帶負電。這使得防禦素像一塊塊小磁鐵,吸附到病毒囊膜帶負電荷的糖蛋白上。這樣便使囊膜病毒穿孔,形成突破口,內容物外泄而死亡。
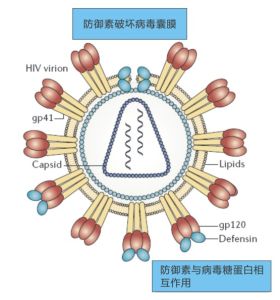 殺滅病毒
殺滅病毒 3、地雷陣----阻止病毒基因複製與轉錄
萬一病毒進入細胞內,防禦素可以與細胞膜表面促腎上腺皮質激素(ACTH),人硫酸肝素糖蛋白(HSPG),低密度脂蛋白受體(LDLR)等相結合,從而啟動G蛋白偶聯型受體的級聯放大反應,進一步激活磷酸激酶C。這些細胞信使作用就像埋伏在胞內的地雷,可以阻止整合到宿主基因組之前的病毒複合體進入細胞核或者阻止病毒的基因轉錄。整合不到宿主基因組的病毒隨後被摧毀。
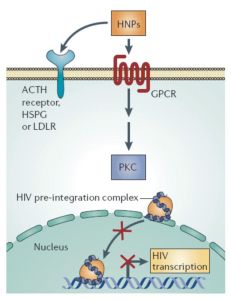 阻止病毒複製
阻止病毒複製 (三) 細胞毒作用
防禦素對正常和惡性哺乳動物細胞都表現出非特異性的細胞毒作用,並且對人淋巴細胞及實體瘤細胞的細胞毒作用更為顯著,尤其對抗TNF的U9TR細胞、抗NK細胞細胞毒因子的小鼠淋巴瘤YAC-1細胞和人組織細胞淋巴瘤U937細胞具有殺傷性。在體外,HNP可抑制煙醯胺腺嘌呤二核苷酸(NADPH)氧化酶的活性,HNP的細胞毒活性部分依賴於靶細胞膜脂質成分的代謝活性,它還可以與過氧化氫酶協同誘導細胞毒作用。體外觀察純化防禦素對人及小鼠數種腫瘤細胞的殺傷作用,發現防禦素對白血病細胞系、淋巴瘤細胞系及實體瘤細胞均具有細胞毒作用,並且這種抗腫瘤功效與其作用時間和劑量呈正相關。
(四)免疫調節作用
防禦素不僅可以直接抵抗病原微生物,而且還具有免疫調節作用。防禦素通過細胞信號傳遞的作用,增強非特異性免疫細胞,尤其是巨噬細胞的活性和趨化性。防禦素還可以促進機體T細胞的趨化和增殖,增強機體免疫應答能力,調節特異性免疫,增強生物機體主動防禦功能。
防禦素能夠作為一種效應分子激活巨噬細胞、DC、氣管上皮細胞等細胞表面受體從而啟動獲得性免疫系統,並將先天性免疫和獲得性免疫有機連線。現已證明一些α-防禦素、β-防禦素對T細胞、單核細胞以及未成熟的DC具有趨化活性,可誘導單核細胞和上皮細胞產生細胞因子。
人、鼠、豬、兔的中性粒細胞防禦素可以誘導肥大細胞脫顆粒並釋放組胺[11]。β-防禦素還能通過與人趨化因子受體6(CCR6)結合,從而吸引不成熟的樹突狀細胞(DC)和記憶T細胞(Tm)至炎症部位,激活細胞免疫和體液免疫。此外,防禦素還可以直接促進感染部位中性粒細胞的補充和積聚。
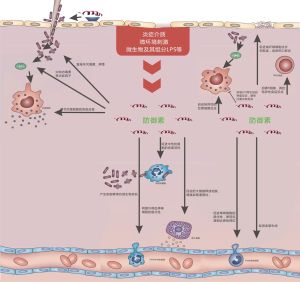 防禦素如何調節免疫系統
防禦素如何調節免疫系統 防禦素的套用
(一)在醫學中的套用
1、防禦素與口腔黏膜疾病的關係
作為機體天然免疫的重要組成部分,防禦素家族在口腔黏膜宿主防禦及免疫應答過程中發揮著重要作用,與多種口腔黏膜疾病(口腔潰瘍、扁平苔蘚、白斑及口腔念珠菌病等)關係密切[5]。現今關於這方面的研究尚處於起步階段,主要以體外實驗為主,其作用機制也多為推測,而有關防禦素在體內的具體作用、有無毒性或其他不良反應等方面的研究甚少,還需進一步全面多層次的研究探討,以闡明防禦素在口腔黏膜疾病發生髮展過程中的具體作用及其機制,為口腔黏膜疾病防治提供新的策略。
2、防禦素與炎症性腸病的關係
克羅恩病(CD)是一種嚴重的腸道慢性炎症性疾病,病變部位可發生於消化道的任何部位,病變形式多樣,其發病是遺傳因素與環境因素共同作用的結果。腸道防禦素是機體抵禦外源性致病因子的重要屏障,而克羅恩病患者腸道防禦素表達水平卻存在著較大差異。迴腸段病變的克羅恩病患者,迴腸潘氏細胞防禦素HD-5、HD-6的表達較正常人和非迴腸段病變克羅恩病患者明顯降低,這種變化與NOD2(nucleotideoligomerizationdomain)基因突變密切相關;結腸組織活檢發現無NOD2基因突變的克羅恩病患者結腸組織HD-5、HD-6的表達水平反而增高,而HD-5、HD-6水平降低將影響腸道黏膜的天然屏障作用,導致黏膜細菌附著引發進一步損害[6]。
以往認為潰瘍性結腸炎(UC)在歐美國家比較多見,但近年來中國文獻報導的病例數也顯著增多。UC病變主要累及結腸,正常人結腸黏膜很少表達HD-5和HD-6,而UC患者結腸上皮細胞中HD-5和HD-6mRNA的表達水平顯著高於正常人,且HD-6的表達還具有特異性,即在非感染性的結腸炎組織中表達並不明顯。HBD-1在腸道上皮細胞呈持續性表達,且不受外界因子影響,具有基礎防禦功能。但套用實時PCR技術檢測發現UC和克羅恩病結腸黏膜組織HBD-1基因表達水平均有所降低,這將可能促使腸道黏膜抗菌活性下降,從而導致細菌入侵機體引發炎症反應[6]。在UC患者的病變結腸組織中,HBD-2可被誘導性表達,而HBD-3的表達水平和HBD-2存在一定的相關性,其在UC時表達亦有明顯增加。此外,HBD-4mRNA的表達水平在UC腸上皮細胞中也明顯增高。
3、防禦素與慢性阻塞性肺部疾病的關係
慢性阻塞性肺部疾病(COPD)是一種以氣道受限為特徵的慢性氣道炎症性疾病,患病率及病死率均較高,其中呼吸道感染是COPD發病及加劇的一個重要因素。HBD-2與呼吸系統疾病關係緊密,在肺部黏膜防禦中具有重要作用。研究證實,HBD-2的產生或作用被抑制後,機體將發生肺部感染[7],據此推測人為補充外源性防禦素可能會阻止細菌的定植和感染的發生。已有研究證實,重組HBD-2對肺損傷具有保護作用,可減少綠膿桿菌感染大鼠肺組織的菌落數[8]。COPD患者存在氣道、肺實質和肺血管炎症,而肺上皮細胞中高表達的HBD-2能通過對肺部炎症因子的調節,阻止全身性炎症反應(SIRS)甚至多器官功能衰竭(MODS)的發生髮展。此外,重組HBD-2還可調控膿毒症誘發的肺組織細胞的過度凋亡,從而減輕肺組織損傷,保護肺功能。
4、防禦素的臨床藥用研究
防禦素抗菌譜廣泛,作用機理特殊,微生物不宜產生抗藥性,因此具有很大的藥用潛力。2008年,全球首個防禦素藥物(PMX-30063)在加拿大完成一期臨床試驗,其對十餘種耐藥菌效果顯著。針對金黃色葡萄球菌引起的急性細菌性皮膚感染這一適應症目前已經完成Ⅱ期臨床研究。它通過直接靶向細菌細胞膜而達到殺菌的目的,不產生耐藥性,從而有望解決日漸增多的細菌耐藥性問題。
(二)在食品工業中的套用
防禦素具有良好的熱穩定性,作為食品防腐劑可用於熱加工食品的防腐保鮮,此外,它還可被用於防止巴氏殺菌後的再污染。
(三)在畜牧業中的套用
抗生素添加劑的使用嚴重破壞了動物腸道的微生態平衡,藥物殘留也影響了畜產品的品質和人類健康,而來源於哺乳動物的防禦素相對分子質量較小,熱穩定性和水溶性均較好,可以在腸道內吸收。由於防禦素屬於多肽成分,在體內容易被蛋白酶降解為胺基酸,動物採食後在體內一般無殘留。利用基因工程方法生產環保型防禦素飼料添加劑或通過日糧添加,都可調控防禦素基因的表達並實現其功效,如減少死胎、木乃伊及殘疾;提高初生重及仔豬出生均勻度[10];提高仔豬斷奶重、斷奶成活率,降低哺乳期仔豬發病率[11];降低哺乳母豬發病率,減少母、仔豬各種應激;改善母豬亞健康和繁殖障礙,延長母豬使用年限;提高保育豬成活率,降低發病率;提高保育豬日增重、降低料肉比;改善免疫抑制,提高免疫應答能力,抗體水平更整齊[12]。
(四)在農業中的套用
防禦素在農業中可用於農作物抗病育種研究,該研究主要是寄希望建立防禦素生物反應器,大量生產並提純防禦素蛋白,以便培育抗性新品種。已有研究者將兔防禦素NP-1轉入小麥植株,田間抗病蟲鑑定結果顯示小麥對於白粉病、葉銹和條鏽病的抗性均有較大提高。而將兔防禦素NP-1基因構建至植物表達載體中獲得轉基因番茄植株的抗病實驗結果表明,其對番茄青枯病具有抗性,為番茄的抗病育種工作奠定了基礎[13]。
六、防禦素的基因工程製備
防禦素主要可通過3條途徑獲取:從細胞或體液中提取、化學合成、重組表達與純化。防禦素在組織中的表達量極少,純化工藝的難度與成本也較高,因此基因工程製備無疑是大量生產防禦素的首選方法[14]。採用先進的基因工程技術將防禦素基因經分子改造後轉化到酵母中進行大規模重組表達與發酵,可以大大提高防禦素的含量和活性,有效地對付病原微生物。
七、防禦素的套用前景
由於傳統抗生素的毒副作用及耐藥菌株的出現,人們正在努力尋找抗生素的替代品。防禦素作為一種新型生物活性肽,其抗菌譜廣泛,與抗生素阻斷大分子生物合成的作用機制完全不同,防禦素能夠快速殺滅廣譜病原微生物,而且作為機體本身的一種活性物質,相對不具有免疫原性,對其具有抵抗性的細菌較少,病原菌也不易對其產生耐藥性,因此可以替代抗生素髮揮廣譜高效的抗菌作用。
隨著生物技術的飛速發展、新防禦素基因的不斷湧現、防禦素家族研究的不斷深入、高效表達系統和生物反應器的不斷更新,我們相信防禦素必將作為新一代抗菌藥物走入我們的生活,為人類創造更多的價值。
八、參考資料
[1] AUVYNETC,ROSENSTEINY.Multifunctionalhostdefensepeptides:Antimicrobialpeptides,thesmallyetbigplayersininnateandadaptiveimmunity[J].FEBSJ,2009,276(22):6497-6508.
[2]李濤,楊銀鳳.防禦素的研究進展[J].內蒙古農業大學學報,2009,30(1):321-324.
[3]高小艷,劉順德,王長法,何洪彬.動物防禦素研究進展[J].家畜生態學報,2010,31(3):80-83.
[4]周春江,黃占景,沈銀柱,等.兔防禦素NP-1基因在小麥抗病育種中套用的初步研究[J].麥類作物學報,2003,23(2):7-10.
[5]LiuWZ,DanHX,MengWX,etal.Relationshipbetweendefensinsandoralmucosaldiseases.InternJOralMed,2008,35Suppl:140-142.(inChinese)劉文釗,但紅霞,孟文霞,等.防禦素與口腔黏膜疾病的關係.國際口腔醫學雜誌,2008,35增刊:140-142.
[6]ZhouL,ZhouCM.Defensinstatusininflammatoryboweldisease.ChinJImmunol,2006,22(12):1161-1164.(inChinese)周聯,周成梅.防禦素在炎症性腸病中的地位.中國免疫學雜誌,2006,22(12):1161-1164.
[7]GanzT.Defensinandhostdefense.Science,1999,286(5439):420-421.
[8]WangHH,ShuQ,ShiZ,etal.Protectiveeffectofrecombioantβ-definsin-2onthelungagainstacuteinjuryinducedbypseudomonasaeruginosainfectioninrats.ChinJAnesthesiol,2005,25(10):762-764.(inChinese)王海宏,舒強,石卓,等.重組β-防禦素-2對呼吸道綠膿桿菌感染大鼠急性肺損傷的保護作用.中華麻醉學雜誌,2005,25(10):762-764.
[9]QiuXY.Antimicrobialpeptidesoftheimmunefunctionandapplicationprospects.BullBiolo,2002,37(1):24-25.(inChinese)邱曉燕.抗菌肽的免疫功能和套用前景.生物學通報,2002,37(1):24-25.
[10]員月明、莫俊、吳指揮、汪猜。防禦素對母豬生產性能及仔豬生產性能的影響[J].養豬,2011,7,10(4)。
[11]曹滿湖、汪猜、肖小平。銀杏防禦素對仔豬生產性能和免疫力的影響[J].動物營養學報,2011,12(6)
[12]肖小平,陳衛富,蔣國成,張維平,吳指揮,堯國福。基因工程防禦素對保育豬生長性能的改善[J].養豬,2011,6.
[13]ZhangXH,GuoDJ,ZhangLM,etal.TheResearchontheExpressionofRabbitDefensin(NP-1)GeneinTransgenicTomato.ActaGenetSin,2000,27(11):953-958.(inChinese)
[14]ZhangSF,CaoRB,JiaY,etal.ModificationofhybridantimicrobialpeptideCecA-milgeneanditsover-secretionexpressioninPichiapastoris.ActaMicrobiologicaSin,2005,45(2):218-222.(inChinese)張素芳,曹瑞兵,賈贇,等.雜合抗菌肽CecAmil的改造及在畢赤酵母中的分泌表達.微生物學報,2005,45(2):218-222.
