
破譯過程
基因密碼的破譯是六十年代分子生物學最輝煌的成就。先後經歷了五十年代的數學推理階段和1961-1965年的實驗研究階段。 1954年,物理學家George Gamov根據在DNA中存在四種核苷酸,在蛋白質中存在二十種胺基酸的對應關係,做出如下數學推理:如果每一個核苷酸為一個胺基酸編碼,只能決定四種胺基酸(4 =4);如果每二個核苷酸為一個胺基酸編碼,可決定16種胺基酸(4 =16)。上述二種情況編碼的胺基酸數小於20種胺基酸,顯然是不可能的。那么如果三個核苷酸為一個胺基酸編碼的,可編64種胺基酸(4 =64);若四個核苷酸編碼一個胺基酸,可編碼256種胺基酸(4 =256),以此類推。Gamov認為只有4^3=64這種關係是理想的,因為在有四種核苷酸條件下,64是能滿足於20種胺基酸編碼的最小數。而4 =256以上。雖能保證20種胺基酸編碼,但不符合生物體在億萬年進化過程中形成的和遵循的經濟原則,因此認為四個以上核苷酸決定一個胺基酸也是不可能的。1961年,Brenner和Grick根據DNA鏈與蛋白質鏈的共線性(colinearity),首先肯定了三個核苷酸的推理。隨後的實驗研究證明上述假想是正確的。
1962年,克里克用T4噬菌體侵染大腸桿菌,發現蛋白質中的胺基酸順序是由相鄰三個核苷酸為一組遺傳密碼來決定的。由於三個核苷酸為一個信息單位,有4 =64種組合,足夠20種胺基酸用了
破譯密碼的競賽中,美國的尼倫伯格博士走在前面。他用嚴密的科學推理對蛋白質合成的情況進行分析。既然核苷酸的排列順序與胺基酸存在對應關係,那么只要知道RNA鏈上鹼基序列,然後由這種鏈去合成蛋白質,不就能知道它們的密碼了嗎?用僅僅含有單一鹼基的尿嘧啶(U),做試管內合成蛋白質的研究。合成蛋白質必須將DNA上的遺傳信息轉錄到RNA上,而RNA的鹼基與DNA稍有不同,一般是有UCGA4種(DNA中是TCGA)。這個實驗只用了含有單一鹼基U的特殊RNA。這樣,就得到了只有UUU編碼的RNA。把這種RNA放到和細胞內相似的溶液里,如果上述觀點正確,應該得到由單一一種胺基酸組成的蛋白質。這樣合成的蛋白質中,只含有苯丙氨酸。於是,人們了解了第一個蛋白質的密碼:UUU對應苯丙氨酸。隨後,又有人用U—G交錯排列合成了半胱氨酸—纈氨酸—半胱氨酸的蛋白質,從而確定了UGU為半胱氨酸的密碼,而GUG為纈氨酸的密碼。這樣,人們不僅證明了遺傳密碼是由3個鹼基排列組成,而且不斷地找出了其他胺基酸的編碼。
進一步研究發現,不論生物簡單到只一個細胞,還是複雜到與人一樣高等,他的遺傳密碼是一樣的。也就是說,一切生物共用一套遺傳密碼。
研究發展
破譯密碼的實驗研究先後由三個實驗逐步發展了四種破譯方法,於1965年完成。
體外實驗
1)在體外無細胞蛋白質合成體系中加入人工合成的polyU 開創了破譯遺傳密碼的先河。
自1961年發現mRNA後,許多實驗室開始在無細胞蛋白質合成系統中加入mRNA,去研究蛋白質生物合成過程,並表明加入mRNA能刺激無細胞系統中蛋白質合成。1961年,美國NIH的Nirenberg和Mathaei,構想:即然mRNA有刺激無細胞系統中的蛋白質合成作用,加入人工合成的多聚核苷酸亦將會有這種促進作用。按此構想,他們合成了polyU作為模板,以觀察無細胞系統中蛋白質合成速率。因為在反應體系中加入高Mg2+濃度,可有利於IF(起始因子)的作用和fMet-tRNANet的形成,從而保證肽鏈合成的起始不需mRNA的適當信號。當把翻譯產物分離、純化和做序列分析後,結果出乎意料,合成的肽鏈中的胺基酸殘基全部是苯丙氨酸,即polyPhe。於是第一次確認了UUU是Phe的密碼子。這樣,就在一個偶然的機會開創了破譯密碼的工作。隨後,他們又以polyA和polyC為模板,證明了分別可指導合成polyLys和polyPro,即確定了AAA是Lys的密碼子,CCC是pro的密碼子。但是類似的實驗不能證明GGG是何種胺基酸的密碼子,因為polyG產生牢固的氫鍵結合,形成三股螺鏇,而不與核糖體結合。
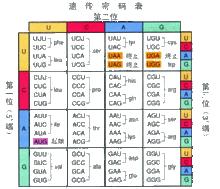 基因密碼錶
基因密碼錶混合共聚物鹼基配對
2)混合共聚物(mixed copolymers)實驗對密碼子中鹼基組成的測定: 1963年,Speyer和Ochoa等發展了用兩個鹼基的共聚物破譯密碼的方法。例如,以A和C原料,合成polyAC。polyAC含有8種不同的密碼子:CCC、CCA、CAA、AAA、AAC、ACC、ACA和CAC。各種密碼子占的比例隨著A和C的不同而不同,例如當A和C的比例等於5:1時,AAA:AAC的比例=5× 5× 5:5× 5× 1=125:25。依次類推。實驗顯示AC共聚物作模板翻譯出的肽鏈由六種胺基酸組成,它們是Asp,His,Thr,Pro,和Lys,其中Pro和Lys的密碼子早先已證明分別是CCC和AAA。根據共聚物成份不同的比例和翻譯產物中胺基酸比例亦不同的關係,Speyer等確定了Asp、Glu和Thr的密碼子含2AlC;His的密碼子含1A2C;Thr的密碼子也可以含1A2C;Pro為3C或1A2C;Lys為3A。但上述方法不能確定A和C的排列方式,而只能顯示密碼子中鹼基組成及組成比例。例如,Asp,Glu和Thr的2A1C可能有三種排列方式,即AAC、ACA、CAA。此外,通過反覆改變共聚物成份比例的方法亦十分麻煩和費時。
aa-tRNA與確定的三核苷酸序列結合
正當Speyer等人按上述2)方法奮力時,Nirenberg和Leder於1964年建立了破譯密碼的新方法,即tRNA與確定密碼子結合實驗。該方法利用了如下事實:即是在缺乏蛋白質合成所需的因子的條件下,特異胺基酸-tRNA(aa-tRNA)也能與核糖體-mRNA複合物結合。最重要的是這種結合併不一定需要長的mRNA分子,而三核苷酸實際上就可以與核糖體結合。例如,當polyU與核糖體混合時,僅有Phe-tRNA(苯丙氨醯-tRNA)與之結合;相應地Pro-tRNA(脯氨醯-tRNA)特異地與polyC結合。還有GUU可促進Val-tRNA(纈氨醯-tRNA)結合,UUG促進Leu-tRNA(亮氨醯-tRNA)結合等。雖然所有64個三核苷酸(密碼子)都可按構想的序列合成,但並不是全部密碼子均能以這種方法決定因為有一些三核苷酸序列與核糖體結合併不象UUU或GUU等那樣有效,以致不能確定它們是否能為特異的胺基酸編碼。
用重複共聚物破譯密碼
4)用重複共聚物(repeating copolymers)破譯密碼:
幾乎在上述Nirenberg和Leder工作的同時,Nishimura,Jones,和Khorana等人套用有機化學和酶學技術,製備了已知的核苷酸重複序列。蛋白質在核糖體上的合成可以在這些有規律的共聚物的任一點開始,並把特異的胺基酸參入肽鏈。例如,重複序列CUCUCUCUCU......是多肽Leu-Ser-Leu-Ser......或者是多肽Ser-Leu-Ser......的信使分子.使用共聚物構成三核苷酸為單位的重複順序,如(AAG)n,它可合成三種類型的多肽:polyLys、polyArg和polyGlu,即AAG是Lys的密碼子,AGA是Arg的密碼子,GAA是Glu的密碼子。又如(AUC)n序列是polyIle、polySer和polyHis的模板。如此至1965年破譯了所有胺基酸的密碼子。
第二套系統
遺傳學的第二套密碼系統(The second genetic code)
以上所述存在於mRNA中的遺傳密碼稱為經典密碼系統或第一套密碼系統。以下所要討論的第二套密碼系統,蘊含於tRNA分子中,這是自1988年5月份以來在分子生物學領域引人注目的新進展。
1)第二套密碼系統的實驗證據-tRNA分子上某些(個)鹼基對能決定tRNA的特異性。 早在70年代初,一些實驗室就觀察到酪氨酸tRNA(tRNATyr)琥珀抑制子在胺基酸接受柄上的突變能使tRNATyr錯誤地攜帶谷氨酸(Glu)。同樣,CUA琥珀抑制反密碼子也能引起其它一些tRNA誤被谷醯化。其後的大約十年,有關方面的實驗證據少有報導。1984年,Prather等發現突變的賴氨酸tRNA(tRNALys)不僅保留對Lys的特異性,而且也能攜帶丙氨酸(Ala)或甘氨酸(Gly)。這個突變的誤義抑制子tRNALys是在胺基酸接受柄螺鏇區的G3 C70被G3 U70鹼基對所取代。能夠使tRNALeu轉變為絲氨酸tRNA(tRNASer)。由此可見,反密碼子在決定tRNA的特異性並非是唯一的關鍵。又比如,琥珀型抑制性半胱氨酸tRNA(tRNACys)苯丙氨酸tRNA(tRNAPhe)和丙氨酸tRNA(tRNAAla),其反密碼子均是CUA,然而它們卻攜帶不同的胺基酸。。colitRNAAla的胺基酸接受柄上單個鹼基對G3 U70,能夠使該tRNA失去負載Ala的功能;進一步將G3 U70引入tRNACys或tRNAPhe,亦可予二者攜帶Ala的功能。他們主要採用上述三種琥珀型抑制性tRNA在二氫尿嘧啶柄(D柄),反密碼柄、TψC柄、胺基酸柄和胺基酸接受柄等部位進行單個或多個鹼基突變,然後檢測宿主E.coliFTP3689的表型抑制作用。發現在總共36種不同突變的tRNA中只有在胺基酸接受柄上A3,C70和C6G7C66G67C70三種突變具有清楚的Sup-表型(即這種宿主E.coly在二天內不生長),而這三種突變共同都有原來的鹼基對G3 U70鹼基對的改變。顯而易見,tRNAAla胺基酸接受柄上G3 U70單個鹼基對決定著Ala的特異性。本文作者也觀察到:只有在多胺下,哺乳動物(大鼠、牛)肝異亮氨酸tRNA(tRNAIle)才能負載ILe。通過測定tRNAIle序列證明它的胺基酸接受柄的G5 G69鹼基不配對。多胺(精胺)通過在此處的橋接,穩定了tRNAIle的空間構象,從而使tRNAIle胺基酸醯化。換言之,即G5 G69對tRNAIle負載可能有決定性作用。
2)第二套密碼系統的概念和特徵
根據上述Hou和Schimmel等人的工作,ChristiandeDuve提出了第二套密碼系統的概念或學說。該學說認為:tRNA胺基酸接受柄有一輔密碼區(Paracodonregion),可以被氨基醯tRNA合成酶(aaRS)識別,並決定tRNA的特異性。他認為第二套密碼系統蘊含於aaRS結構中作者將第二套密碼系統的特徵描述為:[1]與經典密碼系統不同。輔密碼子密碼系統或第二套密碼系統是非簡併性的(nondrgenerate)。可能只有20種aaRS,每種aaRS能夠識別特異於某種胺基酸的所有tRNA,這種識別與該種特異tRNA的不同特徵有關。[2]第二套密碼系統比經典的密碼系統對胺基酸更具有決定性,這與密碼子和相應的胺基酸間的立體化學相互反應有關。認為輔密碼僅與酶-氨基醯-腺苷酸(aaRS-aa-AMP)發生一個非常簡單的反應,而tRNA則起著刪除錯誤氨基醯的作用。[3]第二套密碼系統比經典的密碼系統更原始。一些作者猜測tRNA起源於攜帶氨苷醯的寡核苷酸,其原始形式能與胺基酸直接反應。
3)對第二密碼系統的思考
上文提到的Hou和Schimmel的工作,是ChristiandeDuve提出第二套密碼系統概念的主要依據。在Hou和Schimmel的論文中,只認為在胺基酸柄上的三種突變(A3,C70和C6G7C66G67C70),由於都有原鹼基對G3 U70改變,抑制了tRNAAla的正常負載,沒有觀察到由其它部位突變所產生的影響。tRNA的其它部位對輔密碼子區特異性功能的發揮可能具有協助作用。因為單依靠tRNA分子的單個鹼基對決定其負載的特異性,這不僅尚未得到用其它tRNA大量實驗證實,而在理論上造成tRNA氨基醯化錯誤機率增高,相應使遺傳變異的危險性加大。不論是經典的密碼系統或第二套密碼系統的表達均需aaRS。aaRS與tRNA倒L型構象的內側結構(包括胺基酸接受臂和柄,D反密碼和環)結合。然而,已有的結論認為與aaRS相接觸的tRNA部位對aaRSR的識別作用是非必需的。至今已發表的tRNA序列(除個別低等的生物的外),幾乎都含76或77個鹼基,其中15或16個位點是保守性的(conserved position)即固定地在所有tRNA中存在。這些位點是:U8,A14,G18,G19,A21,U33,G53,T54,ψ,C56,A58,U60,C61,和3'末端的C75,C76,A77。其它61個部位在不同來源和不同tRNA中是可變的(variable position)tRNA上61個可變鹼基與mRNA中含的61個有意義密碼子達到巧合。
然而,deDuve認為第二套密碼系統存在於aaRS結構中,並假定僅僅是輔密碼子與aaRS-aa-AMP或aa-aaRS複合物的一個簡單眼應。酶是一種蛋白質,而蛋白質怎么能作為攜帶遺傳信息的載體呢?aaRS上的某些區域含有一些殘基可與輔密碼子的核苷酸反應,但無法把所有胺基酸側鏈與tRNA的核苷酸匹配起來。一些科學家提出RNA在原始時代具有多種功能,例如攜帶信息,催化活性和轉移信息。在進化過程中,才形成分工負責,即RNA將攜帶信息的功能交給DNA,催化活性由酶(蛋白質)承擔,RNA本身僅保留轉遞信息功能。至今上述三種功能仍不同程度殘留於RNA的事實是對上述分子進化的強有力支持。據此認為:第二套密碼系統存在於tRNA分子本身,而不應存在於aaRS結構中。
破譯展望
但立即受到人們高度注目,破譯第二密碼系統的意義不僅僅限於tRNA分子本身生物學功能的認識,更重要的是將對生物化學,生物起源,分子生物學及遺傳學產生重大影響。
