
神經肌肉接頭
正文
運動神經元軸突末梢在骨骼肌肌纖維上的接觸點。位於脊髓前角和腦幹一些神經核內的運動神經元,向被它們支配的肌肉各發出一根很長的軸突,即神經纖維。這些神經纖維在接近肌細胞,即肌纖維處,各自分出數十或百根以上的分支。一根分支通常只終止於一根肌纖維上,形成1對1的神經肌肉接頭。從神經纖維傳來的信號即通過接頭傳給肌纖維。神經肌肉接頭是一種特化的化學突觸(見突觸),其遞質是乙醯膽鹼(ACh)。無脊椎動物如螯蝦神經肌肉接頭的遞質是谷氨酸(興奮性纖維的遞質)或γ-氨基丁酸(抑制性纖維的遞質)。關於神經肌肉接頭的傳遞是電,還是化學過程,有過很長時期的爭論。奧地利科學家O.勒維在蛙心灌流標本上,首次給突觸的化學傳遞學說以實驗證明,而把這一學說套用到神經肌肉接頭的,則是英國科學家H.H.戴爾等人(1936)。但直到20世紀50年代初微電極技術套用於接頭研究之後,其化學傳遞學說才得到最終確立。
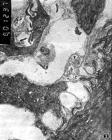 神經肌肉接頭
神經肌肉接頭有些脊椎動物骨骼肌的運動終板不一定是板片狀,如蛙的神經肌肉接頭就是樹枝狀。一般如非特殊指出,終板膜專指屬於肌細胞側的接頭膜,即接頭後膜。神經末梢側的接頭膜叫接頭(或突觸)前膜。接頭前與後膜間存在寬約50納米的間隙──突觸間隙。突觸間隙與細胞外間隙相通,其中充滿細胞外液和散在一些纖維基質。在此纖維基質上附有乙醯膽鹼酯酶。神經末梢的胞漿內含多數線粒體和大量的直徑約50納米的球形小泡──突觸泡。突觸泡內含ACh(據計算約為1萬個ACh分子)。它們在神經末梢內不是平均分布的,而是沿神經末梢長軸每隔約1微米,並在靠近突觸前膜側匯聚成叢。根據突觸泡假說,突觸泡是從此地把內含的遞質ACh釋放到突觸間隙,因而這些突觸泡匯聚的地方叫做活動區。用冰凍蝕刻術製成的接頭標本在電鏡下觀察,可見活動區有一與末梢方向垂直的電子緻密帶,突觸泡在帶的兩側排成單行或雙行,它們可能即是待釋放的突觸泡。在突觸泡的近旁還可見到平行排列的跨膜粒子,有人認為可能是Ca2+通道。終板膜不是平坦的,而是相當有規律地形成許多長約0.7微米,寬約0.8微米的皺褶,叫突觸皺。突觸皺的存在使終板膜面積擴大了約4~5倍。皺褶的嵴部大致與活動區相對應,因而從活動區釋放出的ACh可通過較短距離到達終板膜,與位於其中的乙醯膽鹼受體(AChR)相遇。在突觸皺嵴部分布的AChR密度要比在谷底部的高兩個數量級。
接頭傳遞過程 神經末梢的直徑很小(如人的運動神經末梢的直徑約2~3微米)故傳導動作電位的速度很慢;如在蛙測得的速度為0.4米每秒。當一個神經衝動傳導到神經末梢時,即由它引起去極化,使接頭前膜中的電壓依賴性Ca2+通道開放,Ca2+沿濃度差內流入神經末梢,觸發活動區處的突觸泡與接頭前膜融合併開口,將內含的ACh釋放到突觸間隙(此過程稱胞吐)。據計算一個神經衝動可觸發幾百個突觸泡同步地釋放ACh。釋放出的ACh迅速擴散、通過突觸間隙,到達終板膜,與AChR結合,導致終板膜對Na+與K+的通透性瞬時升高。這種陽離子通透性變化,是由於受體與ACh分子結合後引起了受體分子構型變化,使其離子通道開放造成的。據計算一個突觸泡所釋放的ACh可打開約 2000條受體通道。AChR的離子通道既允許Na+,也允許K+通過。因此,當AChR離子通道開放時Na+沿濃度差內流,K+沿濃度差外流。由它們所攜帶的淨電流使終板膜瞬時去極化。這種去極化叫做終板電位(EPP)。中國神經生理學家馮德培 (1939)是最早發現EPP的科學家之一。當終板電位超過肌細胞的閾值,出現肌細胞動作電位,通過肌細胞內的興奮-收縮耦在線上制,使得肌細胞收縮。釋放出的ACh不論是否與AChR結合,迅速被突觸間隙內的膽鹼酯酶分解,或通過擴散離開突觸間隙。於是 AChR便為接受下次傳遞做好準備。ACh被水解後所生成的膽鹼大部為神經末梢吸收,用於ACh的再合成。這種合成在神經末梢的胞漿內進行。另一方面,多數突觸泡胞吐之後,接頭前膜面積增加,隨之出現前膜的微小內凹再閉合,在胞漿中形成囊泡(此過程稱內吞)。在胞漿中合成的ACh再充填到囊泡中,又形成了可以釋放ACh的突觸泡。
ACh的量子釋放 在正常情況下EPP的振幅明顯超過肌細胞的興奮閾,因此神經興奮引起的EPP都迅速地過渡到肌細胞動作電位。在實驗中為了單獨記錄EPP,往往在溶液中加一定濃度的箭毒,或改變其某種離子濃度(如提高Mg2+濃度或降低Ca2+濃度)等,便可把EPP的振幅降低到肌細胞閾值以下。用細胞內電極從終板區記錄的 EPP可持續達30毫秒以上,迅速上升,緩慢下降的正向電位變化。由於EPP是終板膜所產生的局部電位變化。因此,在終板區記錄的EPP振幅最大,離開終板區迅速衰減。B.卡茨等發現,在終板區進行細胞內記錄時,即使在不受到刺激的安靜狀態,也可記錄到每秒約1次隨機出現的上升快下降慢,但振幅只有約0.5毫伏,持續約20毫秒的正向電變化。除振幅小和“自發”發生之外,這種電變化在形狀、持續時間和對藥物反應等方面均與EPP相似,因而被稱為小終板電位(mEPP)。它們的振幅波動在0.2至0.6毫伏之間,呈常態分配,因而mEPP被認為,是由某固定數目的 ACh分子為單元(稱“量子”)從神經末梢隨機釋放所引起的。一個量子誘發一個mEPP。這種以“量子”為單位的釋放方式叫做量子釋放。以後卡茨等又在對EPP進行了一系列分析的基礎上,提出EPP是在神經衝動的作用下,由多個量子同步釋放所引起的。後來又有工作表明,其他類型的化學突觸的遞質釋放也是量子式的。另一方面用電子顯微鏡觀察(1954)發現,神經肌肉接頭的末梢中含大量直徑為50納米的球形小泡,E.D.P.德·羅伯蒂斯把它們叫做突觸小泡。在此基礎上卡茨又提出了突觸泡假說,認為一個突觸小泡內所含的ACh即為一個量子,因而mEPP便是由單個突觸小泡“自發”釋放所引起的,而EPP則是由多個(100個以上)突觸小泡同步釋放所引起的。量子釋放說雖已得到了較為普遍的承認,但關於突觸泡假說,則尚有分歧,特別是近有人發現了一些難以用該假說解釋的實驗結果,因而又提出ACh是直接由神經末梢胞漿以量子方式釋出的觀點。
參考書目
B.卡茨著,孫以安譯:《神經傳遞介質的釋放》,科學出版社,北京,1979。(B.Katz,The Release of NeuralTransmittersubstances,Charles C.Thomas Publisher Sprinfield,Illinois 1969.)
S.W.Kuffler,J.G.Nicholls,From Neuron to Brain,acellular Approach to the Fuuction of Nervous System, Sinauer Associated Inc., Sunderland,Massachusetts,1976.
