![生物進化[自然科學概念]](/img/8/2ec/nBnauM3XwYDOzYDN2EDO0IDN0UTMyITNykTO0EDMwAjMwUzLxgzLyIzLt92YucmbvRWdo5Cd0FmLxE2LvoDc0RHa.jpg "生物進化[自然科學概念]")
思想發展
古代觀念
古代人們在栽培植物和馴養動物的生產實踐中,積累了關於生物的形態、構造和生活習性的知識,注意到生物機體的變化以及生物與環境的關係,逐步形成了樸素的生物進化思想。古希臘的亞里士多德通過對他那個時代有關動物的知識的系統整理,把540種動物按性狀的異同分為有血的和無血的兩大群,每群之下又分為若干類。他進一步提出生物等級即生物階梯的觀念,認為自然界所有生物形成一個連續的系列,即從植物一直到人逐漸變得完善起來的直線系列;中國戰國時期匯集的《爾雅》一書記載了生物類型的變化;漢初的《淮南子》一書,不僅對動植物作了初步分類,而且提出各類生物是由其原始類型發展而來的。
 生物進化
生物進化近代科學誕生以前,進化思想發展緩慢,當時廣為流行的是神創論和物種不變論。這種觀點直到18世紀仍在生物學中占統治地位,其代表人物是瑞典植物學家C.von林奈(1707~1778)。他所提出的分類系統雖然有助於揭示生物物種之間的歷史聯繫,但他卻把物種看作是上帝創造的不可改變的產物。隨著生產和科學的發展,積累了許多新的與物種不變相矛盾的事實。在大量事實的影響下,甚至像林奈這樣堅定的神創論者,在晚年也不得不承認由於雜交的結果能產生新種。
近代觀念
與林奈的觀點相反,法國學者G.L.L.布豐(1707~1788)相信物種是變化的,現代的動物是少數原始類型的後代。他把有機體與居住環境聯繫起來,認為氣候、食物和人的馴養等因素可引起動物性狀的變異。1809年,另一位法國學者拉馬克(1744~1829)在其《動物學哲學》中,用環境作用的影響、器官的用進廢退和獲得性的遺傳等原理解釋生物進化過程,創立了第一個比較嚴整的進化理論。其意義在於:否定了神創論和物種不變論,奠定了科學生物進化論的基礎,即生物都不是神創造的,而是由更古老的生物進化而來的。
物種起源
1859年,C.R.達爾文發表《物種起源》一書,論證了地球上現存的生物都由共同祖先發展而來,它們之間有親緣關係,並提出自然選擇學說以說明進化的原因,從而創立了科學的進化理論,揭示了生物發展的歷史規律 。
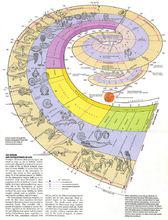 生物進化過程
生物進化過程19世紀80年代以來,以A.魏斯曼(1834~1914)為代表的新達爾文主義把種質論和自然選擇學說相結合,豐富了達爾文的進化理論。20世紀30年代以來,以T.杜布尚斯基(1906~1975)等人為代表的綜合進化論綜合了細胞遺傳學、群體遺傳學以及古生物學等學科的成就,進一步發展了以自然選擇為核心的進化理論。60年代末,日本學者木村資生等人提出中性學說,又在分子水平上揭示了進化的某些特徵,補充、豐富了進化論 。
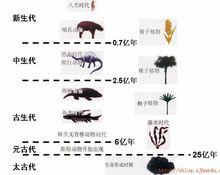 生物進化的一般序列
生物進化的一般序列進化的進步性:地球上的生命,從最原始的無細胞結構狀態進化為有細胞結構的原核生物,從原核生物進化為真核單細胞生物,然後按照不同方向發展,出現了真菌界、植物界和動物界。植物界從藻類到裸蕨植物再到蕨類植物、裸子植物,最後出現了被子植物。動物界從原始鞭毛蟲到多細胞動物,從原始多細胞動物到出現脊索動物,進而演化出高等脊索動物──脊椎動物。脊椎動物中的魚類又演化到兩棲類再到爬行類,從中分化出哺乳類和鳥類,哺乳類中的一支進一步發展為高等智慧生物,這就是人。
主要學說
(1)拉馬克的獲得性狀遺傳學說
(2)達爾文的自然選擇學說
(3)直生論
(4)生物進化的“突變論”
(5)新達爾文主義和綜合進化學說
(6)中性學說
(7)“間斷平衡”學說
(8)進化“四因說”
表1 主要進化理論的比較
| 進化學說 | 生命層次 | 方向 | 動因 | |||||||
| 分子 | 生理 | 個體 | 物種 | 隨機 | 定向 | 定向(趨於完美) | 遺傳 | 生理 | 生態 | |
| 達爾文 | ● | ● | ● | ● | ● | |||||
| 拉馬克 | ● | ● | ● | ● | ● | |||||
| 直生論 | ● | ● | ● | ● | ||||||
| 突變論 | ● | ● | ● | |||||||
| 中性理論 | ● | ● | ● | |||||||
| 間斷平衡 | ● | ● | ● | |||||||
| 四因說 | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● |
進化特徵
 生物進化
生物進化生物界的歷史發展表明,生物進化是從水生到陸生、從簡單到複雜、從低等到高等的過程,從中呈現出一種進步性發展的趨勢。
一般說來,進化過程的進步具有如下特徵:
1. 在生物界的前進運動中,可以看到不同層次的形態結構的逐步複雜化和完善化;與此相應,生理功能也愈益專門化,效能亦逐步增高。
2. 從總體上看,遺傳信息量隨著生物的進化而逐步增加。
3. 內環境調控的不斷完善及對環境分析能力和反應方式的發展,加強了機體對外界環境的自主性,擴大了活動範圍。
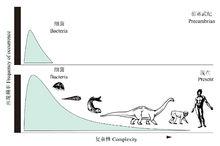 複雜性的進化
複雜性的進化 已經滅絕的愛爾蘭麋鹿
已經滅絕的愛爾蘭麋鹿生物進化的道路是曲折的,表現出種種特殊的複雜情況。除進步性發展外,生物界中還存在特化和退化現象。特化不同於全面的生物學的完善化,它是生物對某種環境條件的特異適應。這種進化方向有利於一個方面的發展卻減少了其他方面的適應性,如馬由多趾演變為適於奔跑的單蹄。當環境條件變化時,高度特化的生物類型往往由於不能適應而滅絕,如愛爾蘭鹿,由於過分發達的角對生存弊多利少,以至終於滅絕 。對寄生或固著生活方式的適應,也可使機體某些器官和生理功能趨向退化。如有一種深海寄生魚,雄體寄生在雌體上,雄體消化器官退化,唯有精巢特別膨大,以保證種族繁衍。
有些研究者對進化的進步性表示懷疑,認為進步性不是進化的基本特徵,也不是進化的本質。科學研究證明,進化不全都引起進步,進化過程中也有退化,但從有機界總的進化過程看,進步性發展是進化的主流和本質。
進化方式
 上海生物進化科普展
上海生物進化科普展生物界各個物種和類群的進化,是通過不同方式進行的。物種形成(小進化)主要有兩種方式:一種是漸進式形成,即由一個種逐漸演變為另一個或多個新種;另一種是爆發式形成,如多倍化種形成,這種方式在有性生殖的動物中很少發生,但在植物的進化中卻相當普遍。世界上約有一半左右的植物種是通過染色體數目的突然改變而產生的多倍體。物類形成(大進化)常常表現為爆發式的進化過程(如寒武紀大爆發),從而使舊的類型和類群被迅速發展起來的新生的類型和類群所替代。
自然選擇
自然界的生物,通過激烈的生存鬥爭,適應者生存下來,不適應者被淘汰掉,這就是自然選擇 。
漸進式
漸進進化是達爾文進化論的一個基本概念,達爾文認為,在生存鬥爭中,由適應的變異逐漸積累就會發展為顯著的變異而導致新種的形成。因為“自然選擇只能通過累積輕微的、連續的、有益的變異而發生作用,所以不能產生巨大的或突然的變化,它只能通過短且慢的步驟發生作用” 。現代進化論堅持達爾文的漸變論思想和自然選擇的創造性作用,強調進化是群體在長時期的遺傳上的變化,認為通過突變(基因突變和染色體畸變)或遺傳重組、選擇、漂變、遷移和隔離等因素的作用,整個群體的基因組成就會發生變化,造成生殖隔離,演變為不同物種。20世紀70年代以來,一些古生物學者根據化石記錄中顯示出的進化間隙,提出間斷平衡學說,代替傳統的漸進觀點。他們認為物種長期處於變化很小的靜態平衡狀態,由於某種原因,這種平衡會突然被打斷,在較短時間內迅速成為新種 。
爆發式
對化石的研究發現,在進化史上,相當長的時間內處於進化較為沉寂的時期,新種的化石很少;有時大量的物種化石集中出現在較短的地質年代,如寒武紀大爆發。
寒武紀是古生代的第一個紀。研究證據表明,寒武紀大約是從距今5.44億年前至距今5.05億年前。地球上最早的生命大約是在距今38億年前出現的。在從38億年前到6億年前這長達32億年的時間裡,生物進化的速率是十分緩慢的。最早的原核生物可能出現在35億年前。最早的真核生物可能出現在20億年前,從那時直至距今6億年前,地球上的生物幾乎都是單細胞生物。從寒武紀開始,地球上突然出現種類繁多的多細胞動物,人們稱這種現象為“寒武紀大爆發(Cambrian explosion)”,也叫“寒武紀大爆炸”。
斷續平衡論
也稱為間斷平衡說。該學說認為化石的不連續性是歷史的真實反映,這正說明生物的進化是不連續的,新物種是短時間內迅速出現的,然後是長時間的進化停滯,直到另一次快速的物種形成出現。
生物的進化既包含有緩慢的漸進,也包含有急劇的躍進;既是連續的,又是間斷的。整個進化過程表現為漸進與躍進、連續與間斷的辯證統一。
進化內容
進化的基本單位
種群是生物生存和生物進化的基本單位, 一個物種中的一個個體是不能長期生存的,物種長期生存的基本單位是種群。一個個體是不可能進化的,生物的進化是通過自然選擇實現的,自然選擇的對象不是個體而是一個群體。 種群也是生物繁殖的基本單位,種群內的個體不是機械地集合在一起,而是彼此可以交配,並通過繁殖將各自的基因傳遞給後代。
進化的原材料
基因庫和基因頻率
基因庫是指一個種群所含的全部基因。每個個體所含有的基因只是種群基因庫中的一個組成部分。每個種群都有它獨特的基因庫,種群中的個體一代一代地死亡,但基因庫卻代代相傳,並在傳遞過程中得到保持和發展。種群越大,基因庫也越大,反之,種群越小基因庫也越小。當種群變得很小時,就有可能失去遺傳的多樣性,從而失去了進化上的優勢而逐漸被淘汰。
基因頻率是指某種基因在某個種群中出現的比例。基因頻率可用抽樣調查的方法來獲得。如果在種群足夠大,沒有基因突變,生存空間和食物都無限的條件下,即沒有生存壓力,種群內個體之間的交配又是隨機的情況下,種群中的基因頻率是不變的。但這種條件在自然狀態下是不存在的,即使在實驗條件下也很難做到。實際情況是由於存在基因突變、基因重組和自然選擇等因素,種群的基因頻率總是在不斷變化的。這種基因頻率變化的方向是由自然選擇決定的。所以生物的進化實質上就是種群基因頻率發生變化的過程。
基因頻率的計算方法
設二倍體生物種群中的染色體的某一座位上有一對等位基因,記作A1和A2。假如種群中被調查的個體有N個,三種類型的基因組成,A1A1、A1A2和A2A2,在被調查對象中所占的個數分別為n1、n2和n3 基因庫和基因頻率的知識可與遺傳的基本規律相結合,在深刻理解遺傳的基本規律的基礎上來理解基因庫和基因頻率的概念就容易得多,也很能夠將這部分知識融會貫通。
基因頻率的改變
引起基因頻率改變的因素主要有三個:選擇、遺傳漂變和遷移。
選擇即環境對變異的選擇,即保存有利變異和淘汰不利變異的過程。選擇的實質是定向地改變群體的基因頻率。 選擇是生物進化和物種形成的主導因素,已經發生的變異能否保留下來繼續進化或成為新物種的基礎必須經過自然選擇的考驗,則自然選擇決定變異類型的生存或淘汰。自然選擇只保留與環境相協調的變異類型(有利變異),可見自然選擇是定向的。經過無數次選擇,使一定區域某物種的有利變異的基因得到加強,不利變異的基因逐漸清除,從而改變了物種在同區域或不同區域內的基因頻率(達爾文只是在個體水平上注意到不同性狀的保留與否,而不能從分子水平對自然選擇的結果加以分析),形成同一區域內物種的新類型或不同區域內同一物種的亞種,或經長期的選擇,使基因頻率的改變達到生殖隔離的程度,便形成新的物種。選擇決定著不同類型變異的命運,也就決定了生物進化與物種形成的方向。
遺傳漂變是指:如果種群太小,含有某基因的個體在種群中的數量又很少的情況下,可能會由於這個個體的突然死亡或沒有交配而使這個基因在這個種群中消失的現象。一般而言,種群越小,遺傳漂變就越顯著。
遷移是指含有某種基因的個體在從一個地區遷移到另一個地區的機會不均等,而導致基因頻率發生改變。如一對等位基因A和a,如果含有A基因的個體比含有a基因的個體更多地遷移到一個新的地區,那么在這個新地區建立的新種群的基因頻率就發生了變化。
可遺傳的變異
可遺傳的變異是生物進化的原始材料,可遺傳的變異主要來自基因突變、基因重組和染色體變異。在生物進化理論中,常將基因突變和染色體變異統稱為突變。基因突變是指DNA分子結構的改變,即基因內部脫氧核苷酸的排列順序發生改變。基因突變是普遍存在的。根據突變發生的條件可分為自然突變和誘發突變兩類。不管在什麼樣的條件下發生突變,都是隨機的,沒有方向性。
染色體變異包括染色體結構的變異和染色體數量的變異,染色體數量的變異又包括個體染色體的增加或減少(非整倍數變化)和成倍地增加或減少(整倍數變化)兩種類型。其中染色體結構的變異與非整倍數變異,由於破壞了生物體內遺傳物質的平衡,所以一般對生物的生命活動是不利的,有時甚至是致命的,在生物進化過程中的意義不大。但染色體整倍數的變化沒有破壞原有遺傳物質的平衡,能夠加強生物體的某些生命活動,對生物的進化,特別是某些新物種的形成有一定的意義如自然界中多倍物種的形成。
基因重組是指染色體間基因的交換和組合。是由於減數分裂過程中,同一個核內染色體複製後發生重組和互換,結果就產生了大量與親本不同的基因組合的配子類型。又由於在有性生殖過程中,雌雄配子的結合是隨機的,進一步增加了後代性狀的變異類型。基因重組實際包括了基因的自由組合定律和基因的連鎖與互換定律。
突變和基因重組都是不定向的,有有利的,也有不利的。但有利和不利不是絕對的,這要取決於環境條件。環境條件改變了,原先有利的變異可能變得不利,而原先不利的變異可能變得有利。等位基因是通過基因突變產生的,並在有性生殖過程中通過基因重組而形成多種多樣的基因型,從而使種群出現大量的可遺傳變異。 變異是不定向的,變異只是給生物進化提供原始材料,不能決定生物進化的方向。
自然決定方向
種群中產生的變異是不定向的,經過長期的自然選擇,其中的不利變異被不斷淘汰,有利變異則逐漸積累,從而使種群的基因頻率發生定向的改變,導致生物朝著一定的方向緩慢地進化。
隔離產生新物種
物種
物種是指分布在一定的自然區域,具有一定的形態結構和生理功能,而且在自然狀態下能夠相互交配和繁殖,能夠產生出可育後代的一群生物個體。
隔離
隔離是指將一個種群分隔成許多個小種群,使彼此不能交配,這樣不同的種群就會向不同的方向發展,就有可能形成不同的物種。隔離常有地理隔離和生殖隔離兩種。
地理隔離是指分布在不同自然區域的種群,由於地理空間上的隔離即使彼此間無法相遇而不能進行基因交流。一定的地理隔離及相應區域的自然選擇,可使分開的小種群朝著不同方向分化,形成各自的基因庫和基因頻率,產生同一物種的不同亞種。分類學上把只有地理隔離的同一物種的幾個種群叫亞種。
生殖隔離是指種群間的個體不能自由交配,或者交配後不能產生出可育的後代的現象。一定的地理隔離有助於亞種的形成,進一步的地理隔離使它們的基因庫和基因頻率繼續朝不同方向發展,形成更大的差異。把這樣的群體和最初的種群放一起,將不發生基因交流,說明它們已經和原來的種群形成了生殖屏障,即生殖隔離。如果只有地理隔離,一旦發生某種地質變化,兩個分開的小種群重新相遇,可以再融合在一起。
地理隔離是物種形成的量變階段,生殖隔離是物種形成的質變時期。只有地理隔離而不形成生殖隔離,只能產生生物新類型或亞種,絕不可能產生新的物種。生殖隔離是物種形成的關鍵,是物種形成的最後階段,是物種間真正的界線。生殖隔離保持了物種間的不可交配性,從而也保證了物種的相對穩定性。生殖隔離分受精前隔離和受精後隔離。教材中提到生物因求偶方式、繁殖期、開花季節、花形態等的不同而不能受精屬於受精前生殖隔離。胚胎髮育早期死亡或產生後代不屬於受精後生殖隔離。
最新成果
新的研究表明,生物在進化過程中,少數幾代能夠從人類導致的不利環境中生存下來,比如氣候變化、過渡捕撈等等。這是因為生物會進化。
瑞典優密歐大學的生物學家托馬斯-卡梅倫表示:“該發現顛覆了人們以前的觀念,即進化的發生速度十分緩慢,往往需要數百甚至上千年的時間。實際上,進化速度很快,往往與生態變化聯繫在一起。”研究人員在實驗室設定了雜亂的生活環境,並捕捉了一些野生土壤蟎類,結果發現它們能迅速地適應生活環境。僅僅經過五代,這些野生蟎就從基因層面進化出了新的特質,扭轉了快要滅絕的勢頭。
卡梅倫表示:“那些具有基因選擇性的蟎類生長速度最為緩慢,但卻擁有極強的繁殖力。因此,我們推測,從長期角度來看,該物種從衰退轉變為興盛的原因是環境對強繁殖力個體的自然選擇作用,當然還包括對子代強適應力的選擇作用。”
科技工作者經常使用蟎類來研究廣泛的生物學疑題,實驗室的發現對人類捕獵動物的管理有著一定的指導作用,如對捕獵管理也許有必要將迅速的進化考慮進去。
