
形態
 阿拉摩龍
阿拉摩龍即使與其他蜥腳類比起來,泰坦巨龍類的頭部顯得比較小。泰坦巨龍類的頭部比較寬,類似圓頂龍與腕龍,不過巨龍類的頭部比較長。它們屬於大鼻龍類,擁有大的鼻孔,而且鼻骨是隆起的,牙齒小型且成鉛筆狀或釘狀。
以蜥腳類而言,泰坦巨龍類的頸部是比較短的。尾巴類似一條鞭子,但沒有梁龍科的尾巴長。泰坦巨龍類的骨盆是比其他蜥腳類纖細,而胸部是比較寬的,讓泰坦巨龍類的四足呈現獨特的步態,因此它們的腳印是比其他蜥腳類來的寬廣。泰坦巨龍類的前肢是比後肢來的粗短,脊椎是實心而非空心的,這也許是一個倒退回原始蜥腳類的特徵。不過它們的脊椎組織是更有彈性,所以泰坦巨龍類可以比其他的蜥腳類移動的更迅速。
從化石的皮膚痕跡發現,在它們皮膚上大的鱗片周圍鑲嵌著較小的鱗片。古生物學家甚至在薩爾塔龍的皮膚上發現類似甲龍的堅甲。
雖然泰坦巨龍類恐龍的體型巨大,但有些物種體型較小,可能是因為異域種化與島嶼環境隔離而造成的侏儒物種,例如馬扎爾龍。
原始泰坦巨龍類具有平凹型或雙凹型脊椎,進階型泰坦巨龍類具有雙凸型脊椎。毒癮龍則具有介於兩者之間的脊椎。
古生物學
食性
在晚白堊紀時期的泰坦巨龍類糞化石中發現了植物岩(Phytoliths)與矽化的植物碎片,顯示了泰坦巨龍類廣泛且不挑剔的食性。一個在2005年公布的發現,顯示泰坦巨龍類有廣泛的進食範圍,除了預期中的蘇鐵與針葉樹,還有單子葉植物,包括棕櫚科、禾本科、稻米與竹的祖先;有假設認為這些新植物造成草食性恐龍與草的協同進化。
蛋巢
最近在阿根廷巴塔哥尼亞的AucaMahuevot發現了一個大型的泰坦巨龍類蛋巢,在西班牙也有類似的發現。很明顯地有數百隻薩爾塔龍科的雌性個體挖掘洞穴,將蛋產於洞中,並用泥土與植被覆蓋恐龍蛋。這些小型恐龍蛋的直徑為11到12厘米,其中包含化石化胚胎,以及完整的皮膚壓痕。皮膚痕跡顯示泰坦巨龍類具有小型的鑲嵌狀鱗片。這些證據顯示泰坦巨龍類是群體動物,身體擁有裝甲,這些裝甲以及群體行動可能用來抵抗大型掠食動物的攻擊,例如:阿貝力龍。
地理與年代範圍
泰坦巨龍類是白堊紀-第三紀滅絕事件之前最晚出現的大群蜥腳下目恐龍,生存於9,000萬年前到6,500萬年前,是當時的優勢草食性動物。化石證據顯示泰坦巨龍類取代其他的蜥腳類恐龍,例如梁龍科與腕龍科,這群動物在晚侏羅紀到中白堊紀期間逐漸被取代、滅亡。
泰坦巨龍類的分布廣泛,尤其是南方各大陸(岡瓦納大陸),甚至在澳洲昆士蘭州也發現屬於泰坦巨龍類的蜥腳類化石。在昆士蘭州內陸小鎮所發現的新標本,身長約為25米長,發現於有9,600萬前歷史的地層,顯示澳洲曾經有大型的泰坦巨龍類生存者。在2006年3月2日有報導傳出,在義大利發現了某種泰坦巨龍類的四個保存良好的骨骸。在2009年,紐西蘭也發現疑似泰坦巨龍類的化石。在2011年,阿根廷科學家宣稱在南極洲曾發現泰坦巨龍類的化石,這代表泰坦巨龍類分布於每個大陸。
分類
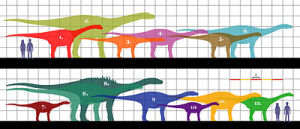 不同泰坦巨龍類與人類的體型比較圖。1.蘇尼特龍、2.澳洲南方龍、3.高橋龍、4.毒癮龍、5.馬拉威龍、6.林孔龍、7.博納巨龍、8.風神龍、9.薩爾塔龍、10.馬扎爾龍、11.博妮塔龍、12.岡瓦納巨龍
不同泰坦巨龍類與人類的體型比較圖。1.蘇尼特龍、2.澳洲南方龍、3.高橋龍、4.毒癮龍、5.馬拉威龍、6.林孔龍、7.博納巨龍、8.風神龍、9.薩爾塔龍、10.馬扎爾龍、11.博妮塔龍、12.岡瓦納巨龍泰坦巨龍類是群分布廣泛而且成功的演化支,但以草食性恐龍而言,泰坦巨龍類的化石紀錄非常少、化石破碎。泰坦巨龍類大約有50個種,但直到最近才發現了頭顱骨與相當完整的骨骸,例如掠食龍。許多物種的所知有限,隨者對於這個演化支的了解增加,許多的化石材料被視為無效,或被重新分類。
泰坦巨龍科是以所知有限的泰坦巨龍屬為名,泰坦巨龍屬是在1877年由理察·萊德克(RichardLydekker)所建立,根據一個部份股骨與兩個不完整背椎。曾經有14個種被歸類於泰坦巨龍屬,使泰坦巨龍屬分布於阿根廷、歐洲、馬達加斯加、印度、以及寮國,並橫跨白堊紀約6,000萬年。儘管泰坦巨龍屬於泰坦巨龍類的分類系統與生物地理學,但對於泰坦巨龍屬所有種的重新審視,使泰坦巨龍屬只剩下5個已承認種。模式種印度泰坦巨龍(T.indicus)是無效種,因為該種原本是根據將廢棄的特徵。因此,泰坦巨龍屬的使用大量地遭到遺棄。泰坦巨龍屬中的最著名標本從此被重新歸類於其他屬,包含伊希斯龍。然而,如果印度泰坦巨龍在未來被重新敘述,根據新的發現,泰坦巨龍屬將暫時性變成有效屬。
有些古生物學家,例如保羅·塞里諾(PaulSereno)的2005年定義,提出泰坦巨龍屬因為資訊過少,因此不能作為定義的基準點,所以不能將其他屬歸類於以泰坦巨龍屬為模式屬的科(例如泰坦巨龍超科、泰坦巨龍科、泰坦巨龍亞科)。Weishampel等人在《TheDinosauria》一書的第二版,也沒有使用泰坦巨龍科,而是使用數個範圍較小的科,例如薩爾塔龍科與納摩蓋吐龍科。
分類學
以下主要根據湯瑪斯·荷茲(ThomasHoltz)等人的2012年研究:泰坦巨龍類Titanosauria阿馬格巨龍Amargatitanis(阿根廷)
安第斯龍Andesaurus(阿根廷)
Atacamatitan(智利)
吉普賽龍Atsinganosaurus(法國)
澳洲南方龍Austrosaurus(澳洲)
俾路支龍Balochisaurus(巴基斯坦)
巴羅莎龍Barrosasaurus(阿根廷)
包魯巨龍Baurutitan(巴西)
布羅希龍Brohisaurus(巴基斯坦)
似傾齒龍Campylodoniscus(阿根廷)
丘布特龍Chubutisaurus(阿根廷)
戈壁巨龍Gobititan(中國)
高橋龍Hypselosaurus(羅馬尼亞)
朱特龍Iuticosaurus(英格蘭)
江山龍Jiangshanosaurus(中國)
九台龍Jiutaisaurus(中國)
卡龍加龍Karongasaurus(馬拉威)
海特蘭龍Khetranisaurus(巴基斯坦)
拉布拉達龍Laplatasaurus(阿根廷、烏拉圭)
利加布龍Ligabuesaurus(阿根廷)
大尾龍Macrurosaurus(英格蘭)
馬拉圭龍Malarguesaurus(阿根廷)
Marisaurus(巴基斯坦)
巴基龍Pakisaurus(巴基斯坦)
沼澤巨龍Paludititan(羅馬尼亞)
普爾塔龍Puertasaurus(阿根廷)
清秀龍Qingxiusaurus(中國)
汝陽龍Ruyangosaurus(中國)
蘇萊曼龍Sulaimanisaurus(巴基斯坦)
怪味龍Tangvayosaurus(寮國)
泰坦巨龍Titanosaurus(印度)
烏貝拉巴巨龍Uberabatitan(巴西)
溫頓巨龍Wintonotitan(澳洲)
真泰坦龍類Eutitanosauria銀龍科Argyrosauridae
風神龍科Aeolosauridae
隆柯龍類Lognkosauria
岩盔龍類Lithostrotia
埃拉爾巨龍Elaltitan
納拉姆布埃納巨龍Narambuenatitan
南極龍科Antarctosauridae
納摩蓋吐龍科Nemegtosauridae
薩爾塔龍科Saltasauridae
系統發生學
在“TheDinosauria”一書的第二版中,泰坦巨龍類被定義為:在蜥腳下目之中,親緣關係較接近薩爾塔龍,而離腕龍較遠的所有物種,這個定義被保羅·阿普徹奇等人的研究(2004年)所沿用。少數科學家,例如保羅·塞里諾,繼續使用節點定義,或是排除盤足龍與腕龍的定義。
在歷史上,泰坦巨龍類的內部關係隨者各種研究而有非常大的不同,因為不同科學家所使用的演化支與等級名稱並不一致。以下為其中一個可能的演化樹,根據Calvo與其同事的2007年研究。他們使用較廣義的泰坦巨龍科範圍,並建立了新演化支,名為隆柯龍類(Lognkosauria)。
新發現化石
 新發現化石
新發現化石2014年5月16日,阿根廷特雷利烏,阿根廷中部丘布特省的特雷利烏市博物館宣布發現了身長超過40米的泰坦巨龍類恐龍化石。之後,該地區又宣布發現200多塊古生物化石,分屬於至少七頭恐龍。這也引發了人們關於該地區是否是一座“恐龍公墓”的猜測。
