")
有孔蟲(化石)
正文
有孔蟲是一類真核單細胞原生生物,除極少數代表如奇桿蟲亞目的部分屬種生活於淡水外,絕大多數生活于海底或營海洋浮游生活。它們個體微小,身體直徑大多小於1毫米,是微體生物,屬於原生生物界肉足動物門根足蟲綱的有孔蟲目,有孔蟲的軟體是由原生質形成的單細胞,分外圍的外質和中部的內質兩部分,外質伸出絲狀或分枝狀偽足,有攝取食物,排除廢物的功能,又是司運動和固著身體的工具;內質包含細胞核及食物泡等,司生殖、營養等各種生活作用。軟體居於殼內,殼由原生質分泌有機質或者有機質加礦物質構成,有些有孔蟲分泌膠結物,再由膠結物粘結海底的砂粒和生物骨骼碎屑構成。有孔蟲個體小、豐度大,具特徵明顯多變而易於保存的殼,地質歷程雖長(從寒武紀延續至今),但各地質時代的發展變化顯著,這些特點使它們成為生物地層學、古生態學和解釋古環境的重要工具和運用數理統計方法進行研究的理想對象。
正式描述的現生有孔蟲大約有6000種,而已知的化石種大大超過此數。在研究和描述現生和化石有孔蟲時都以殼體為主要根據。從細胞學、生活史、行為習性及其他現生生物特徵等角度研究過的現生有孔蟲不足50種。又因為有孔蟲在地質調查及礦產勘探中具有多方面和很高的套用價值。所以它們的發現、描述、分類和其他方面的研究多半是古生物學家的工作。
有孔蟲殼按其成分和構造可分幾類,有機質殼的殼壁呈柔軟有彈性的薄膜狀,統稱為蛋白有機質殼或假幾丁質殼,如奇桿蟲亞目;膠結殼由原生質分泌有機質或礦物質膠結物,粘結海底砂粒,其他礦物顆粒和動物骨骼碎屑形成,礦物質膠結物常為鈣質、鐵質或矽質;矽質殼的殼壁由分泌的蛋白石或二氧化矽構成,如矽房蟲亞目。大多數有孔蟲具鈣質殼,鈣質殼又可分方解石質和文石質兩大類,其超微構造又有多種變化,因類而異。
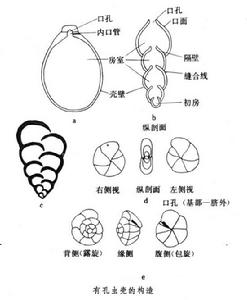
有孔蟲殼的構造從簡單到複雜多種多樣,最基本的構造型式為以下幾種(見圖):
 有孔蟲(化石)
有孔蟲(化石)列式殼由多個房室組成,房室排列成一個系列, 如Nodosaria屬(圖b),第一個房室為初房,分開相鄰兩個房室的隔板叫隔壁,隔壁周邊與殼表交切的一道痕跡叫做縫合線,最末一個房室叫終室,終室向外界的開孔為口孔。殼表口孔周圍的區域叫口面。Nodosaria的房室排成一個系列,有的屬房室排成雙列稱雙列式殼,如Textularia(圖c)屬,也有排列成3列的稱三列式殼,如Fijiella屬。
旋卷式殼由多個房室旋卷排列而成,使殼呈圓餅狀或圓錐狀,以初房為中心,後生的房室沿著初房的對稱面,環繞垂直對稱面的軸盤旋排列,形成平旋式殼,房室每繞軸360°即構成一個旋圈,平旋式殼兩側對稱,如Pu-llenia; 以初房為起點,後生的房室圍繞通過初房中心的軸,螺旋排列,形成螺旋式殼,螺旋式殼兩側不對稱,背側可見初房及所有的旋圈和房室;臍側中央凹陷成臍,只見最後一個旋圈的房室;如Heterolepa(圖e有孔蟲殼還有許多其他類型的構造,一般都由這幾種基本型式演變組合而來,例如Fusulina為兩側包旋的平旋式殼;Bigenerina的殼幼年期雙列式,成年期變為單列式;Cussidulina為旋卷的雙列式殼。一部分粟米蟲類殼的構造獨特,後生的房室環繞初房生長,繞初房一圈生兩個房室,相繼生長的兩個房室的對稱面相交成一定夾角,如Quinqueloculina的夾角為144°,Triloculina的夾角120°,Pyrgo的夾角180°,亦即相繼生長的兩個房室的對稱面在同一平面上。
有孔蟲分兩個生態類群,底棲類群占有孔蟲的大部分,從濱海潮間帶到深海平原都有分布,但在大陸架海底透光帶內最繁盛,它們以絲狀偽足在海底緩慢運動或固著生活,鈣質殼底棲有孔蟲在接近碳酸鹽補償深度(CCD)處逐漸絕跡,超過這個深度仍能生存的是膠結殼有孔蟲,大洋中島山周圍的海底也有底棲有孔蟲分布,底棲有孔蟲的動物群特徵隨生活環境的變化而變化;它們的化石群是解釋陸緣海及其濱海地區的古環境、恢復古地理和研究海岸線變遷的重要標誌。浮游有孔蟲是大洋浮游生物群的重要組分,現代浮游有孔蟲生活在從表層到1000米以上深度的水體中,平均每立方厘米的海水中有10個個體。從赤道到兩極都有浮游有孔蟲分布,其動物群隨著緯度變化的特徵變化明顯,暖流水團與冷流水團中的浮游有孔蟲面貌迥異,因此可以作為研究海洋古氣候及古水文特徵的重要依據。
有孔蟲屬級以上分類單元的劃分和描述,主要以殼體特徵為依據,這些特徵包括殼的一般形態及其變化,內部構造及飾變,口孔形狀及構造、殼壁成分、結晶形狀及分層性,壁孔、管系及莖系的發育情況與微構造等。
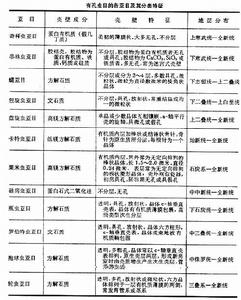
有孔蟲的系統分類研究始於1826年,法國學者A.-C.-V.德薩利納·德奧爾比尼描述了有孔蟲200多個屬,將它們分屬於5個科;至19世紀中葉屬及科的數目增加約一倍,從此,近一個世紀以來,H.B.布雷迪(1884),查普曼(1902),J.A.庫什曼(1927,1933,1940,1948),J.加洛韋(1933),格萊斯納(1945),西格爾(1952),波科爾尼(1954,1958),勞澤爾-切爾諾烏索娃和富爾先科(1959),洛布利奇和H.塔潘(1964),尼古(1979)和海恩斯(1981)先後發表了15部研究有孔蟲系統分類的著作,其中為同行學者普遍接受,套用廣泛者首推J.A.庫什曼(1948)和洛布利奇和H.塔潘(1964)的兩部專著。有孔蟲系統分類的每一次修改都是資料積累增加和研究工具與方法進步的結果,隨著高倍光學顯微鏡的改善,電子顯微鏡的套用和學科間的交叉滲透,使人們對有孔蟲的晶體光學性質與殼壁超微構造有了進一步的認識。洛布利奇與塔潘基於他們的新認識先後於1982和1984年著文對他們自己1964年的分類作了修改補充,1988年發表《有孔蟲屬及其分類》一書,以新的分類觀點,鑑定了3620個屬,確認了其中的2455個,其餘被認為同物異名或因現有信息不足不能成立。並將所確認的屬歸納為12個亞目(見表),74超科,296科和302亞科。
 有孔蟲(化石)
有孔蟲(化石)近年來隨著大洋歷史研究的進展,發現研究有孔蟲化石在論證海底擴張學說方面起著重要作用,最有意義的是,通過對從侏羅紀以來幾類大型底棲有孔蟲從拉丁美洲到中東分布的一致性與分異的研究,為大西洋的擴張史和特提斯海的衰亡史提供了證據;同時,在測定大西洋洋中脊兩側線性磁異常進而推算海底擴張速率的研究中,遇到不能做放射性測年的岩體,則利用其上下直接接觸的沉積層中的有孔蟲組合研究、群落對比以及鈣質殼中氧同位素比率測定其時代。
化石有孔蟲研究最主要是套用於石油天然氣的找礦勘探和海洋地質與海洋歷史研究方面。世界各地海相生儲油地層大多富含有孔蟲化石,有孔蟲化石的研究是隨著石油開發事業的發展而發展起來的,20世紀20年代以來一直主要是為油田地層劃分和對比服務,60年代以後逐漸側重於古生態的研究,為推斷油氣生成與儲集環境及分析環境條件服務。
